Что такое серотониновые рецепторы
Успешное лечение депрессии началось с появлением в 30-е годы электросудорожной терапии. Затем к этому методу добавилась фармакотерапия: в 50-е годы — гетероциклическими антидепрессантами и ингибиторами моноаминоксидазы, в 60-е годы — литием, а в 70-е годы — стабилизирующим настроение противосудорожным средством карбамазепином.
Чтобы ослабить побочные эффекты (например, сухость в носоглотке, запор, электрическую нестабильность миокарда, обмороки, седатацию), были разработаны вещества, повышающие концентрацию серотонина (5-гидрокситриптамина [5-НТ]) в соответствующих синапсах, в том числе флуоксетин.
Вслед за ним быстро появились флувоксамин и сертралин, а затем ряд агонистов и антагонистов серотониновых (5-НТ) рецепторов. Выяснилось, что эти рецепторы делятся на различные типы и подтипы со специфическими функциями.
Серотонин, или 5-НТ, является регуляторным нейромедиатором, приводящим главным образом к тормозным эффектам. Он синтезируется из L-триптофана, который проникает через гематоэнцефалический барьер (сам серотонин к этому не способен), поглощается клетками центральной нервной системы и в них превращается в 5-гидрокситриптамин, т. е. 5-НТ.
Тела нейронов серотонинергической системы расположены прежде всего в шве, или области срединной линии, ствола головного мозга. Они образуют самую крупную сеть с единым нейромедиатором в головном мозге млекопитающих.
Типы серотониновых рецепторов
Известно 4 главных типа серотониновых рецепторов: 5-НТ1, 5-НТ2, 5-НТ3 и 5-НТ4. Первый тип делится на подтипы А, В, С, D и Е, а второй — на подтипы А и В. Рецепторы 5-НТ3 локализованы как в периферической, так и в центральной нервной системе. Антагонисты периферических 5-НТ3-рецепторов, например ондансетрон, гранисетрон и закоприд, применяются для лечения тошноты и рвоты.
Избирательные ингибиторы поглощения серотонина не связываются с какими-либо его специфическими рецепторами, но обеспечивают антидепрессивный эффект, селективно блокируя обратное поглощение этого нейромедиатора пресинаптическими окончаниями, из которых он выделился.

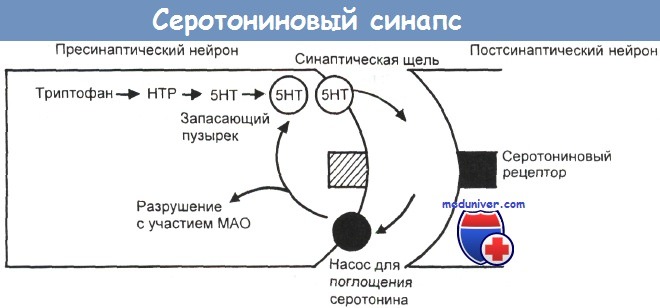 Серотониновый синапс.
Серотониновый синапс.
Серотонин (5-НТ), синтезируемый из триптофана через гидрокситриптофан (НТР), выделяется из пресинаптического нейрона в синаптическую щель.
Попав туда, его молекулы либо действуют на постсинаптический рецептор, обусловливая нервную передачу, либо возвращаются в пресинаптическую клетку с помощью поглощающего механизма насосного типа.
Вернувшись в пресинаптический нейрон, серотонин либо вновь запасается в синаптических пузырьках для будущего высвобождения, либо разлагается моноаминоксидазой (МАО). 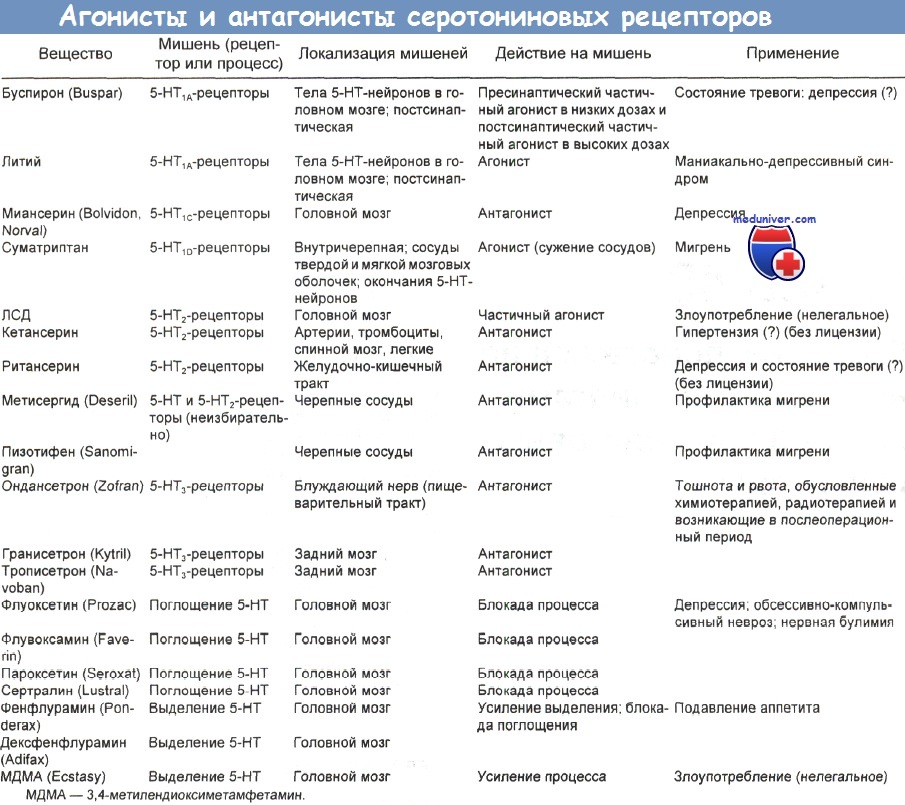

Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Что такое серотониновые рецепторы

Научное название для серотонина 5-гидрокситриптамин или 5-HT. Серотонин в основном обнаруживается в мозге, кишечнике и тромбоцитах.
Серотонин используется для передачи сообщений между нервными клетками, считается, что он участвует в сокращении гладких мышц, и, помимо прочего, способствует положительным эмоциям. Как предшественник мелатонина, он помогает регулировать циклы сна и бодрствования организма и внутренние часы.

Считается, что он играет важную роль в регулировании настроения и моторных, когнитивных и вегетативных функций организма. Тем не менее, точно неизвестно, влияет ли серотонин непосредственно на эти функции, или играет общую роль в координации нервной системы. Вероятно, серотонин участвует в регулировании социального поведения, аппетита и пищеварения, сна, памяти и сексуальной функции. Низкие уровни серотонина связывают с развитием депрессий, однако до сих пор неясно, способствуют ли низкие уровни серотонина депрессии или депрессия вызывает снижение уровня серотонина. Препараты, которые изменяют уровень серотонина, используются для лечения депрессии, тошноты и мигрени, и они могут также играть улучшать состояние при ожирении и болезни Паркинсона. Другие способы повышения уровня серотонина в организме включают в себя повышение настроения, солнечный свет, физические упражнения и диету.

Обычно считается, что серотонин является нейротрансмиттером, хотя некоторые считают его гормоном. Серотонин синтезируется в кишечнике и мозге. Он также присутствует в тромбоцитах и центральной нервной системе (ЦНС). Считается, что он влияет на различные функции организма и психологическое состояние. Серотонин не может проникнуть через гематоэнцефалический барьер.
Как нейромедиатор, серотонин обеспечивает передачу сигналов между нервными клетками (нейронами), регулируя их интенсивность. Считается, что он играет ключевую роль в функционировании ЦНС и организма в целом, особенно желудочно-кишечного тракта. Исследования обнаружили связь между серотонином и метаболизмом в костях, выработкой грудного молока, регенерацией печени и делением клеток. Серотонин влияет на головной мозг. Основная часть серотонина в организме находится в желудочно-кишечном тракте, где он регулирует все его функции, в том числе перистальтику кишечника. Серотонин также играет роль в снижении аппетита во время еды. Серотонин влияет на настроение, уровень тревоги и счастья. Серотонин способствует образованию тромбов. При возникновении повреждения серотонин выходит из тромбоцитов, в результате чего сужаются кровеносные сосуды, уменьшается кровоток и образуются тромбы. В случае попадания в желудочно-кишечный тракт токсичных или раздражающих веществ, кишечник вырабатывает больше серотонина, чтобы увеличить время прохождения пищи и устранить раздражитель. Серотонин также стимулирует рвотный центр в мозге, вызывая тошноту. Некоторые ученые связывают высокий уровень серотонина в костях с увеличением риска развития остеопороза. Серотонин, по-видимому, подавляет сексуальную активность. Селективные ингибиторы обратного захвата серотонина (СИОЗС) повышают уровень серотонина у людей с депрессией, предотвращая реабсорбцию серотонина, что приводит к повышению его уровня в синапсах, но от 20 до 70 процентов людей, принимающих их, испытывают ряд симптомов, связанных с сексуальной дисфункцией.
До настоящего времени не до конца понятно, что именно вызывает депрессию, но основная теория последних 50 лет заключается в том, что причиной депрессии может быть дисбаланс нейромедиаторов или гормонов в организме. Депрессию связывают с низким уровнем серотонина, но остается ли это причиной депрессии или ее следствием, до сих пор неясно.
Тем не менее, ученые в настоящее время ставят под сомнение роль серотонина или любого отдельного нейромедиатора в возникновении депрессии.
При дефиците серотонина наблюдаются ухудшение памяти и плохое настроение. Также низкие уровни серотонина могут привести к тяге к сладкой или мучной пище, плохому сну, снижению самооценки, тревожности и агрессии.
Действительно ли серотонин может помочь при депрессии?
СИОЗС используются с 1980-х годов для лечения депрессии, повышая уровень серотонина. Считается, что такие лекарства, как СИОЗС, облегчают симптомы депрессии, повышая уровень серотонина в организме, но как именно они работают, неясно. Некоторые ученые предположили, что повышение уровня серотонина вряд ли непосредственно улучшит симптомы депрессии. Одна из проблем заключается в том, что можно измерять уровень серотонина в крови, но не в мозге. Исследователи не знают, отражают ли уровни серотонина в кровотоке уровни серотонина в мозге, или могут ли СИОЗС действительно влиять на уровень серотонина мозга. В 2014 году исследование на мышах показало, что серотонин не играет роль в возникновении депрессии. Была создана популяция мышей, в мозге которых не продуцировался серотонин. Эти мыши не проявляли никаких признаков депрессии, даже когда они находились в состоянии стресса.
Однако в 2015 году другие ученые обнаружили, что мыши, у которых не было серотонина, были более восприимчивы к социальным стрессорам, чем здоровые мыши из контрольной группы. Хотя СИОЗС, по-видимому, помогают некоторым людям с депрессией, некоторые ученые теперь утверждают, что «простые биохимические теории, которые связывают низкие уровни серотонина с подавленным настроением, более не являются надежными». СИОЗС также используются для лечения симптомов тревоги, панического расстройства и обсессивно-компульсивных расстройств.
Антисеротонергические лекарственные средства, которые действуют на серотониновые рецепторы, используются для лечения тошноты, вызванной химическими токсинами, включая лекарственные средства, используемые в химиотерапии и общей анестезии.
Серотонинергические вазоконстриктивные противомигреневые препараты или триптаны могут уменьшить симптомы мигрени и хорошо переносятся.
Серотонинергическая система связана с познанием, эмоциями и двигательными функциями. Изменения в этой системе могут влиять на моторные и немоторные функции, обычно связанные с болезнью Паркинсона. В настоящее время исследования в этой области продолжаются.
Повышенная чувствительность к одному из женских гормонов, прогестерону, по-видимому, снижает уровень серотонина в мозге. Ингибиторы серотонина иногда используются для облегчения симптомов предменструального синдрома во время их появления.
Также серотонин может использоваться при лечении ожирения и синдрома раздраженного кишечника.
Серотониновый синдром возникает чаще всего если человек одновременно принимает два препарата, повышающих уровень серотонина, и связан с чрезмерной стимуляцией ЦНС и периферических серотониновых рецепторов. Одновременное использование назначенных врачом лекарств и, например, биологически активных добавок, повышающих уровень серотонина, может привести к развитию серотонинового синдрома. Также развитие серотонинового синдрома возможно при раковых опухолях желудочно-кишечного тракта, поскольку такие опухоли могут вызывать слишком большое выделение серотонина. Признаками серотонинового синдрома являются волнение и беспокойство, спутанное сознание, учащенное сердцебиение и повышение кровяного давления, расширение зрачков, диарея, головные боли, мышечная дрожь, потливость, нарушение координации, ригидность мышц. В тяжелых случаях, опасных для жизни, возможны гипертермия, нерегулярное сердцебиение, судороги и потеря сознания.
Существуют простые и доступные способы повышения уровня серотонина в организме, не связанные с употреблением лекарственных средств. К ним относятся, например, изменение мышления в результате психотерапии или самоиндукции, которые могут повысить уровень серотонина, поскольку взаимодействие между синтезом серотонина и настроением является двусторонним. Физические упражнения оказывают антидепрессивное действие. Использование продуктов, содержащих высокий уровень триптофана, могут улучшить настроение благодаря повышению уровня серотонина.

В ряде исследований пожилые люди, которым давали биологически активные добавки с триптофаном, показали улучшение когнитивных способностей. В настоящее время набирает силу идея о влиянии микробиоты кишечника на ЦНС. В этом случае серотонин в пищеварительной системе может влиять на настроение.
Многое остается неизвестным о серотонине. Трудности, связанные с изучением функций мозга, означают, что пройдет некоторое время, прежде чем можно будет получить полное знание о серотонине.
До сих пор недостаточно доказательств того, что конкретная диета может повлиять на настроение или симптомы депрессии.
Тем не менее, было установлено, что следование здоровой и разнообразной улучшает общее самочувствие. Необходимо сосредоточиться на разнообразном рационе питания и стремиться получать как можно больше питательных веществ из пищевых источников. Применение биологически активных добавок возможно только после консультации с врачом.
Рецептор медиатора настроения
Психонейрофармакологическая важность серотонина столь высока, что его химическая формула даже является узнаваемым образом во многих субкультурах, — в частности, её часто татуируют или исполняют в виде ювелирных украшений.
Автор
Редакторы
В 1948 году, когда был открыт серотонин (5-гидрокситриптамин), никто и не думал, что он настолько важен для физиологии человека [1]. Серотониновая система является молекулярной мишенью при лечении мигреней, депрессии, тревожных расстройств, тошноты, слизистого колита и других заболеваний, своим разнообразием подчёркивающих многогранность роли серотонина в работе всего организма [2]. В некотором смысле серотонин можно считать нейрофармакологической основой духовной сферы человека, — в частности, религиозного и мистического чувств [3].
G-белоксопряженные рецепторы
«Биомолекула» довольно скрупулёзно следит за структурной биологией G-белоксопряженных рецепторов, а поэтому об их роли и истории изучения можно весьма подробно прочесть на нашем сайте:
Рецепторы серотонина являются интегральными белками мембран нейронов, и экспериментальное их изучение затруднено необходимостью использовать мембраномоделирующие среды, позволяющие сохранить природное строение этих белков. Дополнительной хитростью является генно-инженерное встраивание вместо одной из цитоплазматических петель рецептора термостабильного бактериального белка апоцитохрома b562RIL (BRIL), стабилизирующего пространственную структуру и позволяющего формироваться кристаллам дифракционного качества, но не меняющего активность рецепторов. Структуру β2-адренорецептора получили в 2007 году тем же способом, только там вставляли не апоцитохром, а лизоцим, что произвело революцию в структурной биологии GPCR-рецепторов [7].
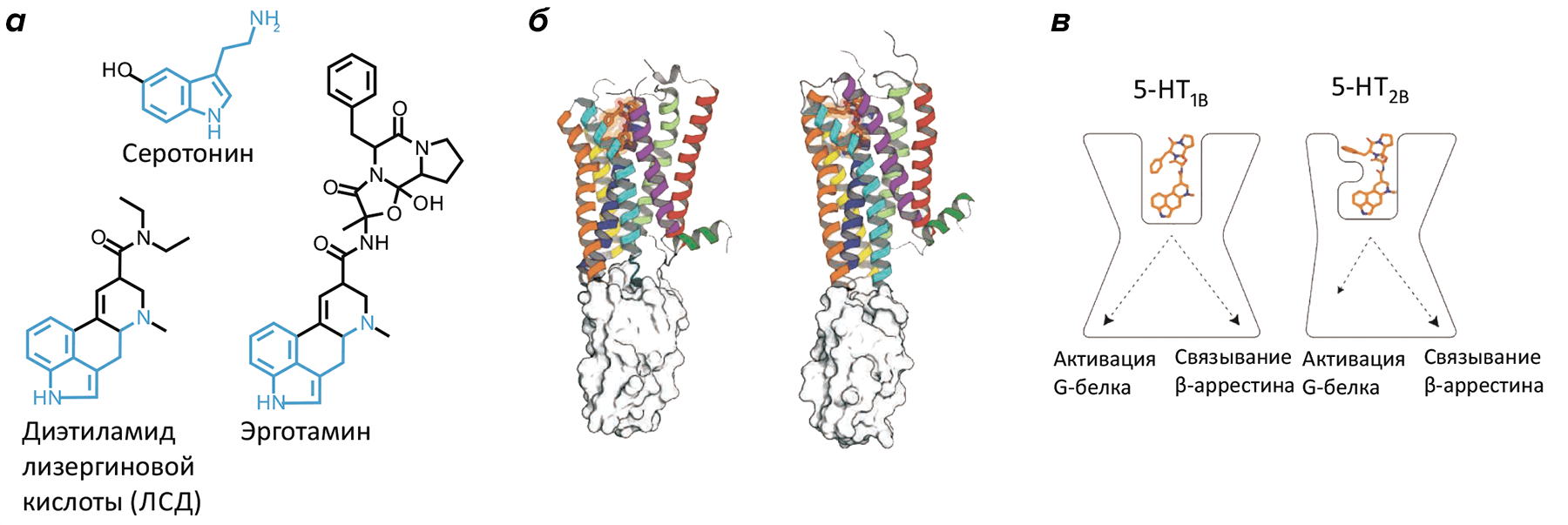
Рисунок 1. Молекулярные основы передачи сигнала серотониновыми рецепторами. а — Природный (серотонин) и синтетические (эрготамин, ЛСД) лиганды с общим индоламинным ядром. б — Кристаллографические структуры рецепторов 5-HT1B (слева) и 5-HT2B (справа) в комплексе с эрготамином. в — Эрготамин стабилизирует различные конформации у двух серотониновых рецепторов, объясняя биохимически установленный феномен «смещенной» рецепторной передачи.
Как уже было сказано, все серотониновые рецепторы, кроме 5-HT3, относятся к GPCR-рецепторам и передают сигнал в клетку либо через активацию G-белка, либо связываясь с альтернативным белком-эффектором — β-аррестином [8]. Близкое химическое строение серотонина и других биологически активных аминов — дофамина и адреналина, — а также высокое сходство рецепторов этих молекул объясняют, почему вещества, действующие на серотониновые рецепторы, могут действовать также на рецепторы других моноаминов, приводя к побочным эффектам. В частности, высокие дозы эрготамина и родственных ему алкалоидов, действующих, предположительно, через 5-HT1B и 5-HT1D рецепторы, могут запустить также α-адренорецепторы, что вызовет нежелательное сосудосуживающее действие [9]. Ещё один довольно скандальный пример связан с фенфлурамином — регулятором аппетита амфетаминовой природы, действующим через рецептор 5-HT2B и приводящим к появлению пороков сердца. Изъятие этого препарата с рынка в 1997 году было крупнейшим в истории фармакологии.
Уже известны примеры рационального дизайна лекарств [10], специфическим образом нацеленных на 5-HT1 рецепторы и используемых для избавления от мигреней [11], однако определение пространственной структуры серотониновых рецепторов обещает сделать процесс этого дизайна более прямолинейным. Детальное сравнение строения двух рецепторов позволит создать вещества, селективно действующие на рецептор 5-HT1B и лишённые кардиотоксического побочного действия, оказываемого через рецепторы 5-HT2B.
На первый взгляд, строение обоих серотониновых рецепторов такое же, как и у наиболее известного представителя семейства А рецепторов GPCR — родопсина [12]. Среднеквадратичное отклонение, используемое в качестве меры схожести двух пространственных структур, здесь составляет всего лишь 2.3–2.7 Å для 80% Cα-атомов любого из серотониновых рецепторов и родопсина, подчеркивая высочайшую похожесть пространственной организации этих рецепторов при совершенно различных функциях и аминокислотных последовательностях. Однако наличие структур двух рецепторов, определенных в комплексе с одним и тем же лигандом, открывает замечательную возможность для тонкого сравнения рецепторных сайтов распознавания и областей, в которых рецептор взаимодействует с цитоплазматическими белками-эффекторами. В частности, удалось объяснить, почему в случае 5-HT2B рецептора эрготамин активирует β-аррестин, в то время как для 5-HT1B это G-белок. Выявление таких тонких молекулярных нюансов — пока редкость для молекулярной биологии, но именно они позволяют предсказывать сигнальный ответ, вызываемый тем или иным соединением, а значит, и прогнозировать и создавать требуемое лекарственное воздействие.
Семейство рецепторов серотонина иногда сравнивают с двуликим Янусом, поскольку одни из них могут быть опасны (5-HT2B называют рецептором смерти из-за его кардиотоксического действия), а другие доставляют нам удовольствие. Например, при поедании шоколада в наш организм поступают предшественник серотонина аминокислота триптофан и некоторые другие серотониноподобные моноамины, и серотониновые рецепторы позволяют нам почувствовать это удовольствие [13].
Знание о деталях строения серотониновых рецепторов, несомненно, найдет применение в дизайне некардиотоксичных аналогов серотонина, которые будут выполнять свою целебную функцию и окажутся приятны во всех отношениях — как шоколад.
По материалам рубрики Perspectives журнала Science [14].
Нейромолекулы: серотониновые рецепторы
Долго у нас не было новых текстов из серии «нейромолекулы». Однако – свершилось. Мы писали много и охотно про нейромедиаторы, «ключах» к дверям синапсов. Однако до сих пор не было ни одной полноценной статьи о «замках» – их рецепторах. Исправляемся, и представляем вашему вниманию большой текст о рецепторах серотонина.


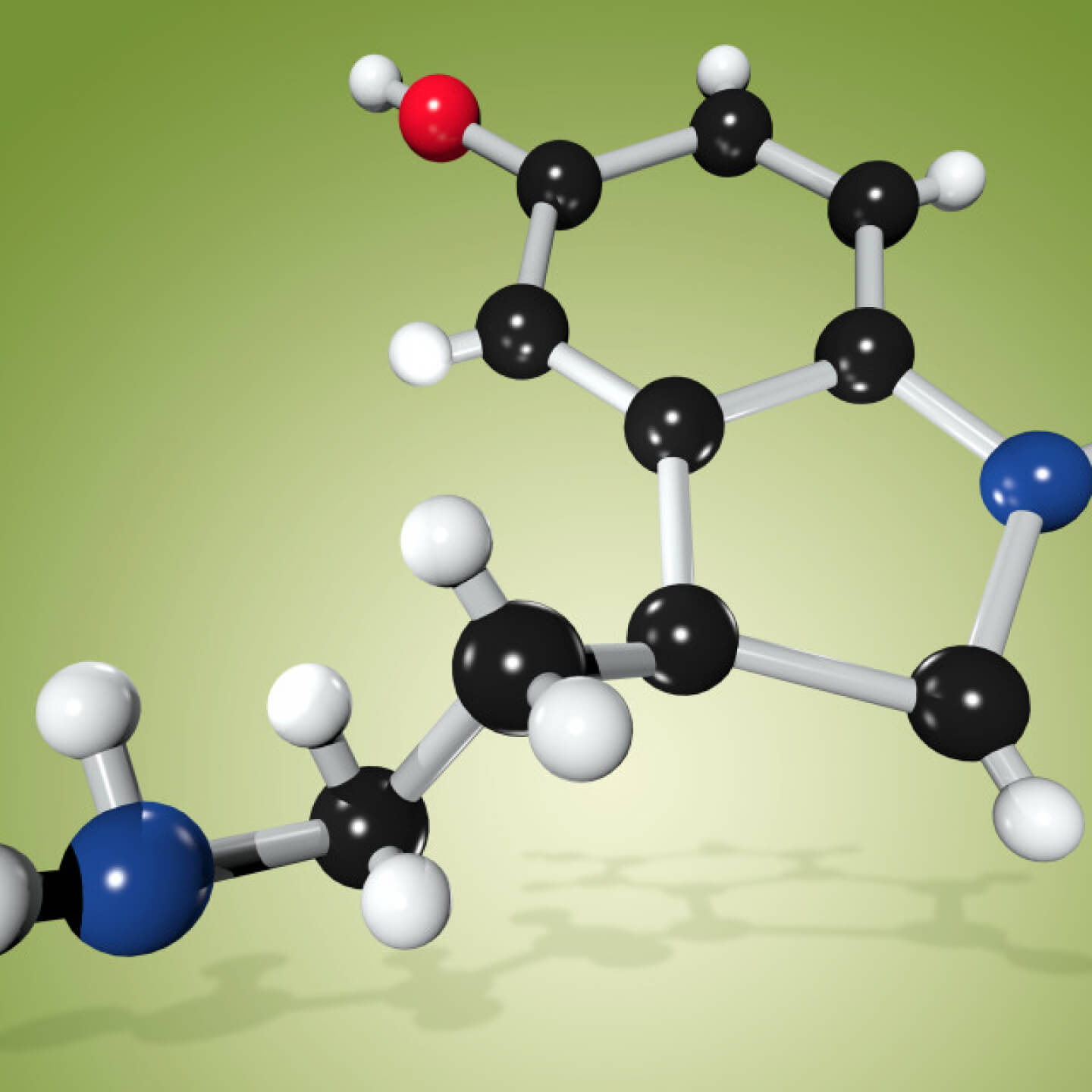
Рис. 1. Химическая формула серотонина (5-гидрокситриптамина)
Основные принципы работы клеточных рецепторов
По принципу действия клеточные рецепторы можно подразделить на две группы: ионотропные и метаботропные. Ионотропные рецепторы, по сути, представляют собой ионные каналы, которые могут открываться или закрываться при связывании с лигандом (то есть небольшой молекулой, которая специфично взаимодействует с рецептором, вызывая изменения в его конформации). Таким образом, ионотропные рецепторы при взаимодействии с лигандом изменяют мембранный потенциал клетки. Метаботропные рецепторы действуют более хитро. После связывания с лигандом они взаимодействуют с особыми мембранными белками — G-белками. Через G-белки опосредованно действуют множество регуляторных молекул организма, например, многие нейромедиаторы и гормоны. Молекулярная биология работы G-белков изучена достаточно подробно и очень сложна, поэтому мы остановимся лишь на некоторых аспектах. G-белки, активированные при взаимодействии с рецептором, запускают внутриклеточные сигнальные каскады, которые, в конечном итоге, изменяют экспрессию генов-мишеней. Источником энергии для G-белков является гуанозинтрифосфат (ГТФ) – молекула, очень похожая на АТФ, но имеющая гуанозин вместо аденозина. В активированном состоянии G-белок связан с ГТФ, однако в процессе работы он гидролизует ГТФ с образованием гуанозиндифосфата (ГДФ) и выключается. В большинстве случаев, после активации G-белки взаимодействуют с одним из двух ферментов: аденилатциклазой или фосфолипазой С. Аденилатциклаза занимается образованием молекул циклического аденозинмонофосфата (цАМФ) – важного вторичного посредника, через который сигнал передаётся дальше вглубь клетки (рис. 1). Фосфолипаза С расщепляет мембранный фосфолипид фосфатидилинозитол-4,5-бисфосфат на два вторичных посредника: диацилгрицерин (DAG), который остаётся в мембране, и инозитол-1,4,5-трисфосфат (IP3), который уходит вглубь клетки, передавая сигнал на всё более высокие уровни.

Рис.2. Схема передачи сигнала через серотониновый рецептор. После связывания серотонина с рецептором происходит активация G-белка, под действием которого аденилаткциклаза превращает АТФ во вторичный посредник – цАМФ. Далее сигнал передаётся вглубь клетки, в конечном счёте приводя к изменению экспрессии генов-мишеней. Источник: http://neuronovosti.ru/neuronbank.org/wiki
Помимо природного лиганда, активирующего рецептор, с рецептором может взаимодействовать множество веществ самой разной природы. Вещества, которые при связывании блокируют работу рецептора, называют антагонистами. Агонисты, как и естественный лиганд, активируют рецептор, причём некоторые из них (обратные антагонисты) изменяют действие рецептора на противоположное.
По месту расположения рецепторов относительно синаптической щели выделяют пресинаптические и постсинаптические рецепторы. Иногда пресинаптические рецепторы ведут себя как постсинаптические, то есть возбуждаются нейромедиаторами, которые выделяет тот же нейрон, на котором находятся эти рецепторы. Такие рецепторы называют ауторецепторами.
Мы разные – в этом наше богатство
Среди рецепторов серотонина есть как ионотропные, так и метаботропные, которые могут действовать как на аденилатциклазу, так и на фосфолипазу С. В наиболее распространённой в настоящее время классификации серотониновых рецепторов (деление на семь семейств) учитываются последовательности и структуры генов и белков, фармакологические свойства и способы дальнейшей передачи сигнала. Дополнительное разнообразие серотониновых рецепторов обеспечивается альтернативным сплайсингом и редактированием их мРНК. В таблице ниже представлены основные свойства разных семейств рецепторов серотонина.
| Семейство | Эффект | Тип | Механизм действия |
| 5-HT1 | Тормозящий | Метаботропный (аденилатциклаза) | Понижение внутриклеточной концентрации цАМФ |
| 5-HT2 | Возбуждающий | Метаботропный (фосфолипаза С) | Повышает внутриклеточную концентрацию IP3 и DAG |
| 5-HT3 | Возбуждающий | Ионотропный Na+/K+-канал | Деполяризация клеточной мембраны |
| 5-HT4 | Возбуждающий | Метаботропный (аденилатциклаза) | Повышение внутриклеточной концентрации цАМФ |
| 5-HT5 | Тормозящий | Метаботропный (аденилатциклаза) | Понижение внутриклеточной концентрации цАМФ |
| 5-HT6 | Возбуждающий | Метаботропный (аденилатциклаза) | Повышение внутриклеточной концентрации цАМФ |
| 5-HT7 | Возбуждающий | Метаботропный (аденилатциклаза) | Повышение внутриклеточной концентрации цАМФ |
Рецепторы серотонина нередко взаимодействуют друг с другом, формируя гетероолигомеры, а также с рецепторами других нейромедиаторов, например, ГАМК [7].
С чем может быть связано такое исключительное разнообразие рецепторов? Серотонин – одна из древнейших сигнальных молекул и, вероятно, его рецепторы прошли свой непростой эволюционный путь уже после того, как они приобрели способность связываться с серотонином. Рецепторы менялись – как структурно, так и функционально, но, несмотря на перемены, они сохраняли способность связываться с серотонином и изменять конформацию при взаимодействии с ним [6].
Ниже мы подробно рассмотрим каждое семейство серотониновых рецепторов.
5-HT1
В этом семействе выделяют пять подтипов: 5-HT1A, 5-HT1B, 5-HT1D, 5-HT1E и 5-HT1F. Практически все они подавляют работу аденилатциклазы, приводя к понижению уровня цАМФ в клетке. Любопытно, что в генах всех пяти подтипов рецепторов полностью отсутствуют интроны, что крайне редко встречается среди генов позвоночных [4] [6].
Наиболее многочисленны рецепторы подтипа 5-HT1A. В N-концевой части молекулы этих рецепторов, находящейся вне клетки, имеются два однонуклеотидных полиморфизма. Один из них ассоциирован с редким нервно-психическим расстройством, известным как синдром Туретта, а второй приводит к утрате чувствительности к серотонину. В центральной нервной системе они обнаруживаются в коре больших полушарий, гиппокампе, прозрачной перегородке, миндалине и ядрах шва, кроме того, небольшие их количества найдены в таламусе и базальных ганглиях. Вне ЦНС они обнаруживаются у нейронов, иннервирующих пищеварительный тракт [4] [6].
Рецепторы 5-HT1A функционируют как ауторецепторы и постсинаптические рецепторы. Они участвуют в регуляции поведения (в том числе пищевого), а также задействованы в развитии тревожности. Фармакологи активно исследуют возможность применения антагонистов этих рецепторов в лечении депрессии и тревожности [4].
Рецепторы 5-HT1B выявляются в ЦНС, где они опосредуют пресинаптическое ингибирование. Они влияют преимущественно на поведение, хотя имеются сведения, что они также регулируют сужение и расширение кровеносных сосудов. С рецепторами этого подтипа взаимодействует эрготамин – алкалоид спорыньи, который действует на них как агонист. Наибольшие количества этих рецепторов обнаруживаются в базальных ганглиях, полосатом теле и фронтальной коре. Функции 5-HT1B-рецепторов зависят от их локализации. В коре они подавляют высвобождение дофамина, а в полосатом теле и базальных ганглиях они функционируют как ауторецепторы и блокируют высвобождение серотонина. Показана их роль в контроле выделения других нейромедиаторов, таких как ацетилхолин, глутамат, дофамин, норадреналин и ГАМК [4]. Любопытно, что при блокировании рецепторов 5-HT1B происходит увеличение количества остеобластов, костной массы и скорости роста костей [8]. Мыши, у которых ген 5-HT1B был выключен, отличаются агрессивностью и тягой к алкоголю [9].
5-HT1D-рецепторы экспрессируются на очень низком уровне. Они функционируют как ауторецепторы в латеральных ядрах шва, однако они также были найдены в сердце, где контролируют выделение серотонина. Рецепторы, находящиеся в ЦНС, участвуют в регуляции движения и тревожности, кроме того, они вызывают сужение сосудов в мозге. Клиническое значение этих рецепторов ещё практически не исследовано. Существуют, однако, лекарства от мигрени, действующие на рецепторы 5-HT1B и5-HT1D [4]. В отличие от рецепторов 5-HT1B, на 5-HT1D эрготамин действует как антагонист.
Рецепторы 5-HT1E практически не исследованы, так как учёные пока не располагают методами очистки, фармакологическими инструментами и животными моделями, которые подходили бы для их изучения. Известно, однако, что гены этих рецепторов одинаковы у разных людей и практически не содержат однонуклеотидных полиморфизмов, поэтому, вероятно, они играют какую-то важную физиологическую роль, пока ещё нам не известную [4].
Рецепторы 5-HT1F представлены в мозге весьма ограниченно, однако они были обнаружены в матке и коронарных артериях. Вероятнее всего, эти рецепторы подавляют аденилатциклазу, однако детального исследования их функций ещё не проводилось [4].
5-HT2
Семейство 5-HT2 серотониновых рецепторов делится на три подтипа: 5-HT2A, 5-HT2B и 5-HT2C (рис. 3). Они представляют собой метаботропные рецепторы, действующие на фосфолипазу С и приводящие к усилению гидролиза фосфатидилинозитол-4,5-бисфосфата и повышению внутриклеточной концентрации кальция. Рецепторы этого семейства – важнейшие возбуждающие серотониновые рецепторы, хотя рецепторы 5-HT2A могут обладать тормозящим действием в некоторых зонах коры больших полушарий [4].
Рецепторы 5-HT2A экспрессируются в самых разных тканях и органах. В частности, они участвуют в сокращении гладких мышц. В ЦНС этот подтип представлен в основном в коре, ограде и базальных ганглиях. При активации 5-HT2A усиливается секреция ряда гормонов, в числе которых АКТГ, кортикостерон, окситоцин, ренин и пролактин, а ингибирование этих рецепторов сказывается на поведении. Разнообразные антагонисты 5-HT2A (например, популярный нейролептик кветиапин) используются или рассматриваются как потенциальные препараты, показанные при шизофрении. Наряду с 5-HT2C, 5-HT2A играют важнейшую роль в поведенческом ответе на галлюциногены [4]. Интересно, что рецепторы 5-HT2A, по-видимому, необходимы для проникновения внутрь клеток вируса Джона Каннингема. Этот вирус проникает в олигодендроциты, астроциты, B-лимфоциты и клетки почечного эпителия, вызывая прогрессирующую мультифокальную лейкоэнцефалопатию [10].
Активация 5-HT2B приводит к сокращению гладкой мускулатуры верхней части желудка. Эти рецепторы обнаружены в мозжечке, латеральной прозрачной перегородке, гипоталамусе и средней части миндалины. Антагонисты 5-HT2B только начинают разрабатываться и, возможно, они найдут применение в лечении и предотвращении мигрени. Есть сведения, что рецепторы этого типа каким-то образом задействованы в регенерации печени [4].
Функции рецепторов 5-HT2C исследованы мало из-за отсутствия селективных лигандов. Антагонист 5-HT2C, известный как агомелатин, является эффективным антидепрессантом, так как повышает уровни дофамина и норадреналина в некоторых отделах мозга. Флуоксетин неспецифично стимулирует рецепторы 5-HT2C за счёт повышения концентрации серотонина в синапсе. Некоторые атипичные антипсихотические препараты частично блокируют эти рецепторы [4]. Ген 5-HT2C располагается на X-хромосоме, которая у мужчин имеется в единственном варианте, а у женщин в каждой клетке одна из двух Х-хромосом инактивируется случайным образом, поэтому влияние однонуклеотидных полиморфизмов в этом гене на структуру и функции белка может быть различным.

5-HT3

Рис. 4. Пространственная структура 5-HT3 рецептора мыши, вид сверху. Разные субъединицы покрашены разными цветами. В центре канала видна пора. Структура получена методом криоэлектронной микроскопии. Изображение из Protein Data Bank (PDB ID: 6BE1)
5-HT4
Известно семь подтипов рецепторов этого семейства (5-HT4A-H), которые различаются своими С-концевыми частями. Тем не менее, все эти рецепторы фармакологически схожи и действуют на аденилатциклазу. Рецепторы 5-HT4 проявляют фоновую активность даже в отсутствие лиганда (то есть серотонина). Из-за этой особенности оценивать эффекты агонистов и антагонистов 5-HT4 очень непросто. Некоторые подтипы демонстрируют тканеспецифичность, например, 5-HT4D встречается только в кишечнике. Ряд подтипов влияет не только на аденилатциклазу, но также на калиевые каналы и потенциалзависимые кальциевые каналы. Функции 5-HT4-рецепторов очень разнообразны. Они влияют на моторику кишечника, сердечные сокращения, в ЦНС регулируют высвобождение серотонина и других нейромедиаторов (ацетилхолина, дофамина и ГАМК), а также усиливают синаптическую передачу, что может сказываться на работе памяти. Один из агонистов 5-HT4-рецепторов, цисаприд, некоторое время использовался в клинической практике для усиления моторики кишечника, но в настоящий момент его не применяют из-за негативного влияния на сердце. Частичный агонист этих рецепторов, тегасерод, начинают использовать как симптоматическое средство при раздражении толстой кишки. Разнообразные селективные лиганды рецепторов 5-HT4 рассматривают как потенциальные средства, эффективные при лечении самых разных заболеваний – артмий, нейродегенеративных заболеваний, недержании мочи. Однако использование сильных и высокоспецифичных агонистов 5-HT4-рецепторов сильно ограничено выраженными побочными эффектами на сердечно-сосудистую систему [4].
5-HT5
У грызунов имеется два подтипа рецепторов этого семейства: 5-HT5A и 5-HT5B. У человека, однако, в гене 5-HT5B имеется преждевременный стоп-кодон, делающий ген нефункциональным, поэтому в мозге человека экспрессируется только подтип 5-HT5A. Фармакологические особенности этих рецепторов практически не изучены. На основании локализации этих рецепторов в различных отделах мозга предполагается, что они могут участвовать в контроле движений и питания, развитии тревожности и депрессии, обучении, консолидации памяти, адаптивном поведении и развитии мозга. Они также могут быть задействованы в регуляции нейронами работы астроцитов. Нарушение этих взаимодействий наблюдается при ряде серьёзных патологий ЦНС, например, при болезни Альцгеймера, а также у больных синдромом Дауна [4].
5-HT6
У человека описаны два варианта рецептора 5-HT6. Первый подтип располагается преимущественно в лимбической и экстрапирамидной зонах коры, а второй – в хвостатом ядре и чёрной субстанции. Клиническое значение этих рецепторов до конца неясно. Возможно, они каким-то образом опосредуют действие некоторых антидепрессантов и антипсихотических препаратов. Имеются свидетельства их участия в холинергической передаче сигнала [4]. Антагонисты 5-HT6-рецепторов могут использоваться для лечения депрессии и нарушений памяти, а также ожирения, поскольку они уменьшают аппетит и вызывают снижение массы тела [11] [12].
5-HT7
У человека рецепторы 5-HT7 представлены несколькими изоформами (5-HT7A-D) и активируют аденилатциклазу, а также MAP-киназу. В центральной нервной системе 5-HT7-рецепторы наиболее многочисленны в таламусе, гипоталамусе, гиппокампе и коре. Эти рецепторы также обильно экспрессируются в сосудах и, по-видимому, участвуют в их расширении. Рецепторы 5-HT7 задействованы в регуляции сна, циркадных ритмах, обучении и памяти [13]. При остром (но не хроническом) стрессе количество 5-HT7-рецепторов увеличивается, а при длительном приёме антидепрессантов, напротив, уменьшается. Антагонисты этих рецепторов могут быть полезны при лечении депрессии и расстройств сна [4].
Мы вместе – в этом наша сила
Из описанного выше колоссального разнообразия серотониновых рецепторов может показаться, что их ничего не объединяет, кроме сродства к серотонину. По счастью (особенно для фармакологов), это совершенно не так. Почти все серотониновые рецепторы – метаботропные и почти все активируют аденилатциклазу. Таким образом, нисходящие эффекты у них принципиально похожи. Кроме того, структуры многих из них очень похожи – не только у рецепторов одного семейства, но и у рецепторов разных семейств. Благодаря этому существует немало соединений, неспецифичных в отношении серотониновых рецепторов и взаимодействующих почти со всеми из них. Скорее, специфичные агонисты и антагонисты серотониновых рецепторов – большая редкость.
Действие большинства антидепрессантов так или иначе связано с серотонином, и многие из них взаимодействуют с серотониновыми рецепторами с разной степенью сродства. Некоторые антидепрессанты связываются только с одним видом серотониновых рецепторов. Например, первый нетрициклический антидепрессант – тразодон – из всех серотониновых рецепторов взаимодействует только с 5-HT2А, действуя на него как агонист [2]. Однако гораздо чаще мишенями антидепрессантов являются разные серотониновые рецепторы. Так, амитриптилин – один из наиболее сильнодействующих антидепрессантов, введённый в клиническую практику ещё в 1961 году, — является антагонистом или агонистом рецепторов 5-HT2A, 5-HT2C, 5-HT3, 5-HT6 и 5-HT7 [3]. Даже самые современные препараты бывают неспецифичны в отношении серотониновых рецепторов. Например, вортиоксетин, который был одобрен для клинического применения всего лишь в 2013 году в США и в 2014 году в Европе, действует как агонист рецепторов 5-HT1A, частичный агонист 5-HT1B и антагонист 5-HT1D, 5-HT3 и 5-HT7 [1].
В заключение можно отметить, что, хотя в наших знаниях относительно серотониновых рецепторов много белых пятен, многие препараты, применяющиеся в психиатрии, взаимодействуют с различными серотониновыми рецепторами – иногда блокируя их работу, иногда усиливая, а иногда вообще меняя их эффект на противоположный. Разработка новых, высокоспецифичных препаратов, обладающих малыми побочными эффектами, — безусловно, важнейшее направление в современной фармакологии, поэтому и детальное изучение серотониновых рецепторов сейчас важно как никогда.
Распространение серотониновых рецепторов в органическом мире
По-видимому, серотонин стал использоваться живыми организмами в качестве сигнальной молекулы очень давно, и его рецепторы обнаруживаются не только у позвоночных и других животных, но даже у растений и грибов [6].
Вероятнее всего, первые серотониновые рецепторы появились 700-800 миллионов лет назад у одноклеточных эукариот вроде инфузории-туфельки. От предковых рецепторов первыми отделились семейства 5-HT1, 5-HT2 и 5-HT6 примерно 750 миллионов лет назад. Рецепторы этих семейств имеются у самых разных животных: от червей (планарий и нематод) до насекомых и позвоночных. 650-700 миллионов лет назад от 5-HT1 отделились два других семейства: 5-HT5 и 5-HT7. Таким образом, большая часть эволюционного пути серотониновых рецепторов произошла ещё до разделения животных на первичноротых и вторичноротых (600-650 миллионов лет назад), поэтому значительная доля серотониновых рецепторов у всех животных примерно одинакова. Однако после расхождения первичноротых и вторичноротых эволюция серотониновых рецепторов в виде дупликаций генов и их дальнейшей специализации продолжилась независимо у этих двух групп животных [6].