Эволюционно мхи представляют тупиковую (слепую) ветвь развития растений, во многом близки к водорослям. Для размножения большинству мхов нужна вода, так как мужские половые клетки могут добраться до женских только с ее помощью. Мы рассмотрим этот отдел на примере кукушкина льна, который относится к подклассу Зеленые мхи.
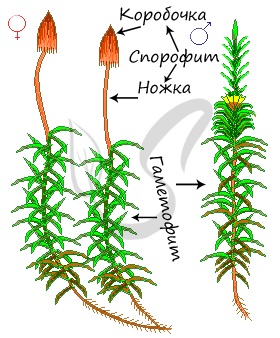
Строение кукушкина льна
Проясним термин «двудомные», указанный вначале. Он означает, что женские (архегонии) и мужские (антеридии) половые органы развиваются на разных растениях. На женском гаметофите расположен архегоний, имеющий бутыльчатую форму, на мужском гаметофите развиваются антеридии. Антеридии окружают характерные крупные листья желто-бурого цвета, по которым всегда можно отличить мужской гаметофит от женского: на женском подобные листья отсутствуют.
В сухую погоду, когда споры созревают, колпачок, а за ним и крышечка, отпадают. Тонкий стебелек спорогона раскачивается от ветра, споры высыпаются.
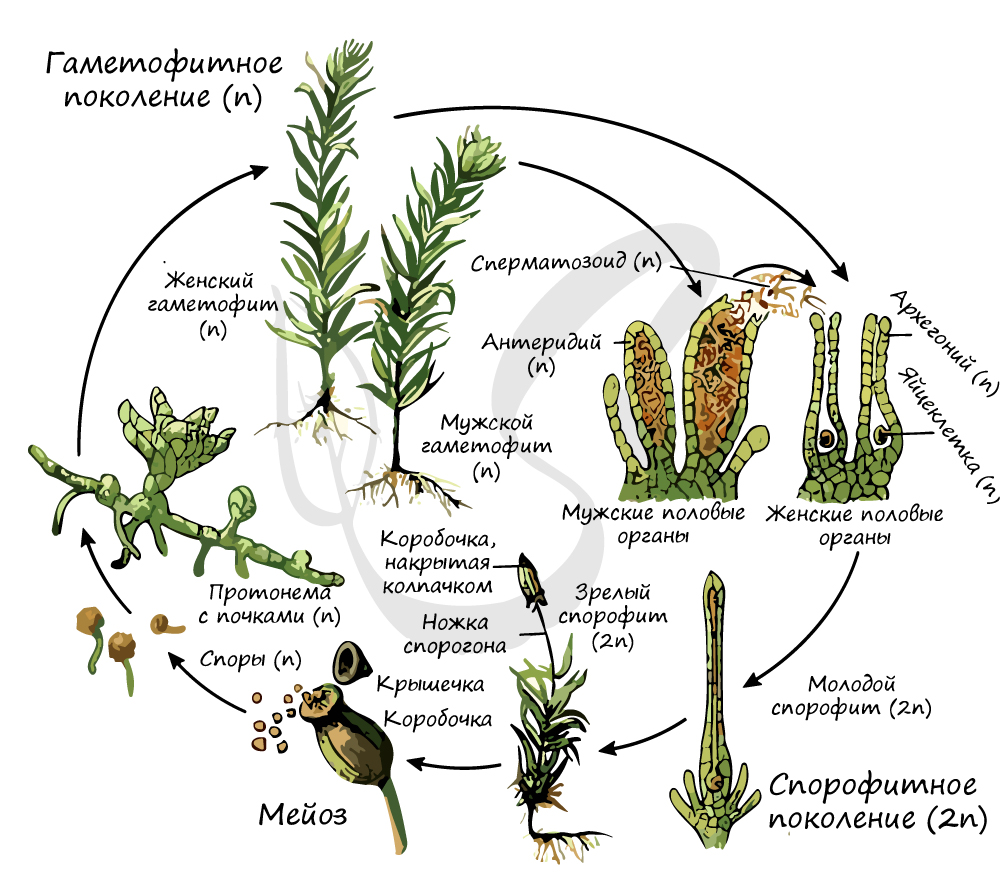
Жизненный цикл мхов
Теперь попробуйте самостоятельно назвать все стадии жизненного цикла мха на картинке ниже, почувствуйте себя истинным бриологом 😉
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
К какому отделу относятся мхи
Ранее в этот отдел, помимо класса листостебельных мхов, или собственно мхов (в ранге класса — Bryopsida), включали также Печёночные мхи (в ранге класса — Marchantiopsida, или Hepaticopsida) и Антоцеротовые мхи (в ранге класса — Anthocerotopsida), но сейчас эти таксоны повышены до уровня собственных отделов Marchantiophyta и Anthocerotophyta. Для обозначения совокупности этих трёх отделов используется неформальный собирательный термин мохообразные (Bryophytes).
Раздел ботаники, предметом изучения которого являются мохообразные, называется бриологией.
Связанные понятия
Упоминания в литературе
Связанные понятия (продолжение)
Неолектомице́ты (лат. Neolectomycetes) — класс грибов отдела Аскомицеты (Ascomycota), один из 4 классов, относящихся к подотделу Taphrinomycotina. Класс монотипный, к нему относятся также монотипные порядок и семейство, соответственно, Neolectales и Neolectaceae, и род Neolecta с четырьмя видами.
Расте́ния-поду́шки, или подушкообра́зные растения, или растения подушкови́дной фо́рмы — жизненная форма растительных организмов, для которой характерны многочисленные короткие, интенсивно ветвящиеся побеги, без явно выраженного главного ствола, между которыми накапливается субстрат состоящий из отмерших частей растения и характеризующийся своеобразной, круглой, подушкообразной формой. Годичный прирост всех побегов одинаков и ничтожен по величине. Принадлежат к различным семействам и родам (азорелла.
У термина Цикадовые есть и другое значение.Саго́вники (устар.: саго́вики, цика́довые) — древняя группа семенных растений, шире всего представленная в мезозое, ныне рассматриваемая в ранге отдела Саговникови́дные (Cycadophyta), в который входит единственный современный класс Саговниковые (Cycadopsida) и единственный порядок Саговниковые (лат. Cycadales, Cycadeae), состоящий из двух семейств.
К какому отделу относятся мхи
Моховидные, или Мхи, или Настоящие мхи, или Бриофиты (лат. Bryophyta ) — отдел высших растений, насчитывающий около 10 000 видов, объединённых примерно в 700 родов и 110—120 семейств [1] (общее число всех мохообразных, включая Печёночные мхи и Антоцеротовые мхи, составляет около 20 000 видов [2] ). Как правило, это мелкие растения, длина которых лишь изредка превышает 50 мм; исключение составляют водные мхи, некоторые из которых имеют длину более полуметра, и эпифиты, которые могут быть ещё более длинными. Моховидные, как и другие Мохообразные, отличаются от других высших растений тем, что в их жизненном цикле гаплоидный гаметофит преобладает над диплоидным спорофитом.
Раздел ботаники, предметом изучения которого являются мохообразные, называется бриологией.
Содержание
История
Многие данные указывают на то, что мхи, как и другие споровые растения, произошли от псилофитов (риниофитов) — группы древних вымерших наземных растений. При этом спорофит мхов рассматривается как конечный результат редукции разветвлённого спорофита предковых форм.
Биологическое описание
У мхов нет цветков, корней и проводящей системы. Размножаются мхи спорами, которые созревают в спорангиях на спорофите. В жизненном цикле, в отличие от сосудистых растений, преобладает гаплоидный (то есть с одинарным набором непарных хромосом) гаметофит (половое поколение). Гаметофит мхов — многолетнее зелёное растение, нередко с листоподобными боковыми выростами и корнеподобными выростами (ризоидами), в то время как спорофит (или бесполая стадия жизненного цикла) короткоживущий, быстро усыхает и состоит только из ножки и коробочки, в которой созревают споры.
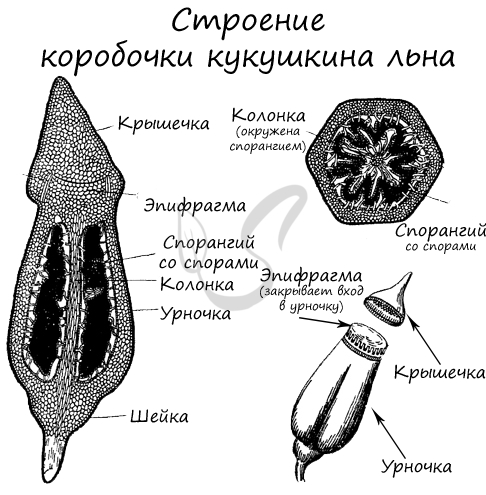
Спорофит моховидных (носящий название спорогоний, или спорогон), имеет более простое строение, чем у других групп высших растений. Он не способен укореняться и располагается на гаметофите. Спорофит состоит, как правило, из трёх элементов:
Со мхами не следует путать так называемый Олений мох (Ягель), поскольку олений мох — собирательное название для нескольких лишайников.
Распространение
Мхи встречаются на всех континентах, в том числе в Антарктиде, нередко в экстремальных условиях обитания.
Обычно мхи образуют плотные скопления в затенённых местах, нередко в непосредственной близости от воды, хотя могут встречаться и на относительно открытых, сухих участках. Среди моховидных есть виды, живущие в пресноводных водоёмах. Морских обитателей нет, хотя некоторые виды поселяются на скалах в полосе прибоя.
Значение мхов
В деятельности человека:
Классы
См. также
Примечания
Литература
Ссылки
Полезное
Смотреть что такое «Мхи» в других словарях:
мхи — (моховидные), отдел высших растений. Включает 22 27 тыс. видов. Выделяют антоцеротовые мхи, печёночные мхи и листостебельные мхи. Известны с карбона. Распространены повсеместно. Особое значение имеют в тундре, где играют ландшафтную роль. В… … Биологический энциклопедический словарь
мхи — (моховидные, бриофиты), отдел высших растений. Преимущественно многолетние растения, характеризующиеся групповыми формами роста (дерновинки, куртинки, подушки). Органы полового размножения архегонии и антеридии, бесполого спорогон (диплоидный) … Энциклопедический словарь
МХИ — МХИ, представители около 14 000 видов мелких, простых по строению нецветковых растений, для которых типично образование обширных колоний, часто образующих плотные ковры. Они размножаются спорами, которые созревают в коробочках на концах длинных… … Научно-технический энциклопедический словарь
МХИ — МХИ, отдел высших растений. Преимущественно многолетние растения, характеризующиеся групповыми формами роста (дерновины, куртины, подушки). Около 20 тыс. видов, по всему земному шару. Нередко способствуют заболачиванию почв, ухудшению качества… … Современная энциклопедия
МХИ — (моховидные бриофиты), отдел высших растений. Преимущественно многолетние растения, характеризующиеся групповыми формами роста (дерновинки, куртинки, подушки). Органы полового размножения архегонии и антеридии, бесполого спорогон (диплоидный),… … Большой Энциклопедический словарь
Мхи — мн. Топкое место, где во множестве растет мох. Толковый словарь Ефремовой. Т. Ф. Ефремова. 2000 … Современный толковый словарь русского языка Ефремовой
МХИ — в широком понимании растения, соответствующие типу мохообразных. Иногда термин понимается более узко: только как класс листостебельных мхов (luusci). Геологический словарь: в 2 х томах. М.: Недра. Под редакцией К. Н. Паффенгольца и др.. 1978 … Геологическая энциклопедия
МХИ — (Bryophyta), класс архегониальных растений (около 18 000 видов), разделяемый на два подкласса: 1) лиственные М. (Musci) и 2) печоночные М. (Hepaticae). Первые имеют стебель, нарастающий верхушкой, и сидящие на нем плоские листья; корни… … Большая медицинская энциклопедия
МХИ — монохроматическое излучение Словарь: Словарь сокращений и аббревиатур армии и спецслужб. Сост. А. А. Щелоков. М.: ООО «Издательство АСТ», ЗАО «Издательский дом Гелеос», 2003. 318 с … Словарь сокращений и аббревиатур
мхи — мхи, моховидные (Bryophyta), отдел высших растений. Преимущественно наземные, многолетние, низкорослые растения. Высотой от 1 мм до нескольких см, реже до 60 см и более. Тело М. гаметофит (половое поколение) представлено слоевищем или… … Сельское хозяйство. Большой энциклопедический словарь
Вон там украдкой слабый луч Скользит по липе, мхом одетой. (А. К. Толстой)
Наиболее древние мхи известны с карбона (по некоторым сведениями – даже с девона) [вообще-то, если 420 млн лет назад, то с силура]. В настоящее время отдел моховидных, или мохообразных (Bryophita) включает в себя около 25 000 видов, разделённых на три класса: лиственные мхи, печёночные (печёночники), антоцеротовые. Иногда антоцеротовые мхи относят к печеночникам, а лиственные мхи называют просто мхами.
Разделы страницы о мохообразных растениях:
Выход растений на сушу
Завоевание растениями суши произошло 420 миллионов лет назад. По-видимому, первыми организмами, заселившими сушу, стали потомки зелёных водорослей. Выход на сушу – это качественный скачок в эволюции, типичный пример аромогенеза (последовательности ароморфозов), потребовавший от растений преодолеть следующие трудности:
Устройство мхов
Все вышеназванные приспособления мы можем наблюдать у современных мхов – наиболее примитивных организмов из группы высших [наземных] растений. Моховидные (Bryophyta) – это отдел царства растений, объединяющий многоклеточные, просто устроенные растения, у которых отсутствует проводящая ткань (флоэма либо ксилема). Именно с последним фактом связаны небольшие размеры этих растений. Тело представителей этого отдела невелико (1–50 см) и слегка дифференцировано на условные «стебель» и «листья», хотя настоящих листьев и стеблей у них нет. Нет у мхов и корней, «стебель» прикрепляется к земле ризоидами – нитевидными выростами стебля, состоящими из одной или нескольких клеток (настоящие корни, в отличие от ризоидов, содержат клетки проводящей ткани).
Мхи достаточно плохо приспособлены к жизни на суше. Они растут, в основном, в сырых и затенённых местах, нередко на гниющей древесине или под водой. Мхи образуют основной растительный покров тундры и торфяных болот. Из-за почти полного отсутствия кутикулы поверхность таллома интенсивно испаряет воду; её недостаток восполняется за счёт поглощения всей поверхностью тела. Некоторые моховидные, однако, могут выдерживать продолжительные периоды засухи, используя какие-то не совсем понятные механизмы. Другие могут расти на открытых скалах.
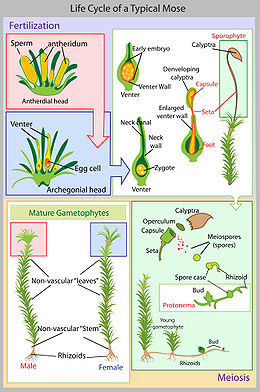
Как и у всех высших растений, у мхов наблюдается чередование поколений. В течение одного жизненного цикла гаплоидный гаметофит в антеридиях образует сперматозоиды, которые с током воды по таллому достигают архегония и сливаются с яйцеклеткой в диплоидную зиготу. Зигота вырастает в диплоидный спорофит – поколение, размножающееся бесполым путём. В результате мейоза на спорофите появляются гаплоидные споры, из которых прорастают гаплоидные гаметофиты, и всё повторяется вновь. У мхов гаметофит является доминирующим поколением, а спорофит, развиваясь и питаясь за счёт гаметофита, полностью зависит от него. Споры образуются в споровой коробочке, возвышающейся над гаметофитом на тонкой ножке.
Так называемые «листья» и «стебли» мхов не являются настоящими стеблями и листьями не только из-за отсутствия сложного тканевого строения, но и потому, что развиваются на другой стадии жизненного цикла – на гаметофите, преобладающей стадии жизненного цикла мхов, а не на спорофитной стадии, как у сосудистых растений.
У мхов также хорошо развито вегетативное размножение при помощи специализированных почек и клубеньков. Почти каждая вегетативная клетка, если её изолировать от прочих, способна вырасти в самостоятельное растение.
Обзор мохообразных
Бриофиты — чрезвычайно своеобразные растения, замечательные во многих отношениях, можно сказать, собрание уникумов. Изучением их занимается особый раздел ботаники — бриология.
В отличие от покрытосеменных растений и от грибов, мохообразные известны (за пределами сверхузкого круга специалистов-бриологов) крайне мало. Даже ботаники, не говоря уже о представителях других биологических дисциплин, проявляют порой неосведомленность, когда речь заходит о мохообразных. Да и трудно ожидать другого. Научно-популярная литература старательно обходит эту группу растений, в учебниках по ботанике она освещается очень кратко и, к сожалению, не всегда на основе современных данных.
Между тем интерес, проявляемый к этой группе растений, с каждым годом возрастает. И далеко не одних ботаников привлекают мохообразные. И даже не только биологов. Это понятно. Бриология в последние десятилетия развивается довольно интенсивно, сведения о бриофитах накапливаются, и становится всё более очевидным, что эти растения заслуживают самого пристального внимания. И не только в плане чисто научном, что в общем-то всегда было ясно и никаких сомнений не вызывало, главное — в плане их практического значения и использования.
Мохообразные обладают многими признаками, которые переводят их из разряда растений, имеющих в основном лишь научный интерес и очень ограниченное практическое значение, в разряд растении, безусловно и многогранно полезных и ценных. Не настолько, конечно, ценных, как покрытосеменные, но все же.
Из живущих на Земле высших растений мохообразные наиболее примитивны. И хотя они только боковая, слепая ветвь эволюции и не являются ничьими предками, без знания мохообразных не решить многих важных вопросов, связанных с изучением всего сонма высших растений, имеющих огромное значение в жизни человека.
Мохообразные малы по размерам и очень нетребовательны. Лабораторная культура их довольно проста и необременительна. Они — удобнейший объект для самых различных экспериментальных исследований, в том числе генетических, и вполне заслужили титул «растительной дрозофилы»1. Именно при работе с мохообразными были получены (впервые в растительном мире) искусственные полиплоиды и открыты половые хромосомы у растений.
В то же время мохообразные — мощные сорбенты. Некоторые из них способны поглотить количество воды, превышающее их собственный воздушно-сухой вес в 20—25 и даже 35 раз! Они не только извлекают из субстрата, на котором растут, различные химические элементы, но и способны поглощать их непосредственно из воздуха, если в субстрате их нет. Это делает мохообразных наряду с лишайниками идеальными (эпитет использован в одной из бриологических работ, и это не преувеличение) индикаторами загрязнения атмосферы.
Жизненный цикл мохообразных
Жизненный путь, пли, как говорят ботаники, жизненный цикл, большинства растений складывается из двух, иногда равных, но большей частью неравных «половинок» — фаз развития, или ядерных фаз, последовательно сменяющих друг друга. Раньше эти фазы именовали поколениями или генерациями.
Растение с двойным (диплоидным) набором хромосом (его называют спорофазой или спорофитом) сменяется растением с простым — гаплоидным — набором хромосом (это растение именуют гаметофазой или гаметофитом). А оно в свою очередь снова сменяется первым. И так далее. Первое растение (спорофит) производит органы бесполого размножения — споры; второе (гаметофит) — органы полового размножения. Их обобщенное название у растений — гаметангии. В результате оплодотворения число хромосом удваивается, и возникает диплоидный спорофит.
У очень небольшого числа водорослей спорофит и гаметофит неразличимы. Но у большинства низших растений и у всех высших (к которым принадлежат и мохообразные) гаметофит и спорофит очень мало или вовсе не похожи друг на друга. Если не знать жизненного цикла растения, то его спорофазу и гаметофазу можно легко принять за совершенно разные, не родственные виды. И это не раз случалось в истории ботаники.
Поведение гаметофита и спорофита в жизненном цикле различных групп растений различно. У некоторых обе фазы существуют отдельно одна от другой, у других — вместе, и при этом одна фаза частично или полностью живет за счет другой.
У папоротников и некоторых других высших растений гаметофит живет отдельно от спорофита, так сказать, сам по себе, а у покрытосеменных и голосеменных — на спорофите (точнее, внутри его).
Не будет преувеличением сказать, что высшие растения осуществили спорофитную, диплоидную линию эволюции. В их жизненном цикле спорофаза резко преобладает.
Но нет правил без исключений. Таким исключением и являются мохообразные. Они пошли в эволюции своей дорогой. У них все наоборот по сравнению с остальными высшими растениями. Если у всех высших само растение, как мы уже говорили, спорофит, то у мохообразных — гаметофит, гаметофаза с гаплоидным набором хромосом. Спорофит у мохообразных имеет меньшие размеры, лишен листьев и по сути дела низведен до роли спороносящего органа. Он прикреплен к гаметофиту и живет в значительной мере за его счет. Еще совсем недавно считалось, что спорофит мохообразных паразитирует на гаметофите. Проскальзывают подобные высказывания порой и в современной литературе. Это, однако, не вполне верно, так как выяснилось, что спорофит мохообразных, по крайней мере в молодом возрасте, фотосинтезирует и, следовательно, о паразитизме, во всяком случае полном паразитизме, говорить нельзя. Нередко считают, что спорофит произрастает на гаметофите мхов эпифитно [за счёт прикрепления], а не как паразит. Правильнее, вероятно, будет сказать, что обе фазы образуют сложный двуединый организм с главенствующей ролью гаметофазы, организм, в котором спорофаза представляет собой спороносный орган. Такие взгляды высказывал известный советский бриолог А. С. Лазаренко [1961]. Сводить сложные и не до конца выясненные взаимоотношения фаз к паразитизму или эпифитизму одной из фаз вряд ли правильно. Добавим, что у части мохообразных (правда, у очень небольшой части) спорофит способен к самостоятельному существованию без какой-либо помощи со стороны гаметофита.
И размеры, и срок жизни спорофита у мохообразных (за очень редкими исключениями) значительно уступают размерам и срокам жизни гаметофита. У очень и очень многих видов мохообразных спорофит возникает даже не каждый год и далеко не на каждом гаметофите. Более или менее регулярно (именно более или менее) спорофиты образуются максимум у 40% видов мохообразных.
Очень любопытно, что, несмотря на кардинальные различия между гаметофитом мохообразных и спорофитом остальных высших растений, между тем и другим поразительно много сходства и, с другой стороны, поразительно мало сходства между гаметофитом мохообразных и гаметофитом остальных высших растений, равным образом как и между спорофитами обеих групп растений. Самое удивительное и неожиданное, что на стебель и листья расчленены спорофиты всех высших растений (кроме мохообразных) и гаметофиты (а не спорофиты!) мохообразных.
Поскольку в одном случае листья образуются на диплоидном растении, а в другом — на гаплоидном, то совершенно очевидно, что сходство этих листьев в большей мере внешнее, чем основанное на общем или одинаковом происхождении. Возникло даже мнение, что лист мохообразных не должен называться листом. Для него были придуманы специальные термины — «филлоид» или «филлид». Точно так же поступили и со стеблем мохообразных — его назвали осью или каулидом.
И это правильно. Разные органы, хотя и похожие, должны называться по-разному. Другое дело, что в повседневной практике термины «лист» и «стебель» в применении к листьям и стеблям мохообразных вполне исправно служат, не создают никакой путаницы. Вот почему термины «филлоид», «филлид», «ось» и «каулид» не получили широкого распространения, оставшись на страницах чисто теоретических исследований. Но помнить о том, что стебель и листья мохообразных принадлежат гаметофиту, в то время как у всех остальных высших растений — спорофиту, надо всегда.
В силу многих причин, касаться которых пока не будем, гаплоидная линия эволюции, избранная и развитая мохообразными, была невыгодной. Остались отрезанными многие возможные пути эволюционного развития. Прежде всего, мохообразные обречены оставаться мелкими растениями. Это лилипуты растительного мира. Самые крупные из них — австралийские мхи из рода Dawsonia (главным образом, Dawsonia superba) и широко распространенный водный мох Fontinalis antipyretica — не достигают одного метра в длину (или в высоту). Их максимальные размеры не более 70 сантиметров. И это — гиганты среди мохообразных.
С малыми размерами связано и то обстоятельство, о котором упоминает советский ботаник Б. М. Козо-Полянский [1965, с. 72]: «В отличие от папоротникообразных и других типов, мохообразные никогда в истории Земли не играли ведущей роли в растительном покрове: эра мохообразных в истории Земли неизвестна». Поэтому другие растения за мохообразными «не пошли». Они избрали иную — спорофитную (диплоидную) — линию эволюции. Последователей у бриофитов не оказалось. Впрочем, почти не было у них и предшественников. Лишь небольшое число водорослей попробовало пойти тем же путем, что впоследствии был пройден мохообразными. Но в воде, как и на суше, этот путь оказался бесперспективным, и основная масса водорослей по нему не пошла.
В итоге мохообразные — единственный отдел растительного царства, в жизненном цикле представителей которого полно и безраздельно господствует гаметофаза.
Чем своеобразнее, необычнее какое-либо явление, тем легче его охарактеризовать. Мохообразные, как мы видели, побили все рекорды по степени своеобразия. И потому они могут быть охарактеризованы предельно кратко: это высшие растения, в жизненном цикле которых преобладает гаметофаза.
Порталы и энциклопедии о мхах
О мохообразных растениях в сети: сборники обзоров, статей, новостей.
Листостебельные (Bryophytina?) мхи (Musci) насчитывают около 15 000 видов (по некоторым данным, до 25 000), объединённых в 660 родов. Преобладают в тундре, на болотах и во влажных лесах. Андреевые мхи имеют мелкие красно-бурые стебли с однослойными листьями. Обычно растут подушечками на скалах высоко в горах.
Гаметофит имеет стебель и листья – обычно однослойные, расположенные на растении по спирали. Ризоиды многоклеточные ветвистые. Сфагновые (торфяные) мхи имеют крупные беловато-зелёные, жёлтые, бурые или красноватые стебли с листьями из чередующихся хлорофиллоносных и воздухоносных клеток.
Лиственные мхи подразделяются на подклассы сфагновых (торфяных), андреевых (створчатоплодных) и бриевых мхов (просто мхов). Последние наиболее распространены в природе.
Подкласс Бриевые, или просто мхи — Bryidae
Бриевые мхи отличаются от прочих механизмом рассеивания спор из коробочки (при помощи специальных зубцов), растут они, как правило, во влажных местах. Активно развиваясь, мхи способствуют заболачиванию почв, ухудшают качество лугов.
Подкласс Сфагновые, или торфяные мхи, сфагны — Sphagnidae
Торфяные (сфагновые) мхи выделяют гуминовые кислоты, препятствующие гниению, благодаря чему скопления мёртвого мха со временем спрессовываются в торф. Торф добывают как полезное ископаемое и используют как топливо или удобрение. Кроме того, из него получают некоторые химические вещества.
Сфагновые мхи используются в качестве подстилки для скота. Так как сфагнум поглощает большие количества воды (в 30–40 раз больше массы своего тела), в войну его использовали как перевязочное средство.
Подкласс Андреевые, или створчатоплодные мхи — Andreaeidae
Печёночные мхи (печёночники; Hepaticae) составляют несколько порядков, около 60 семейств, 280 родов и 9500 видов. Распространены в странах тропического или умеренно влажного климата.
Гаметофит – уплощенное образование, большинство видов имеют «стебель» с расположенными рядами вдоль него листьями. Ризоиды одноклеточные. При рассеивании спор коробочка раскрывается на четыре створки; рассеиванию помогают специальные пружинки – элатеры.
Раньше печёночные мхи делили на два подкласса: маршанциевые и юнгерманниевые печёночники [основные подклассы]. Теперь выделяют 7 групп печёночников:
Антоцеротовые мхи (Anthocerotopsida?) – своеобразная группа, близкая к маршанциевым печёночникам; иногда рассматривается как подкласс печёночных мхов. 2 семейства, 6 родов, свыше 300 видов. От других классов мхов отличаются тем, что в жизненном цикле преобладает стадия спорофита. Слоевище лопастное или розетковидное. Распространены в странах тропического или умеренно влажного климата.
Свое название антоцеротовые получили от греческих слов anthos — цветок и keros — рог: на их розетковидном темно-зеленом пластинчатом слоевище, плотно прилегающем к почве, образуются удлиненные и слегка изогнутые спорогоны роговидной формы. Сравнительно тонкие слоевища антоцеротовых, диаметром 1—3 см, имеют розетковидную или (реже) лентовидную форму и различаются между собой, главным образом, степенью изрезанности (или волнистости) краев. Своеобразный облик придают им довольно многочисленные щетинковидно-торчащие спорогоны высотой до 2—3 см. При созревании спорогоны, вначале зеленые, сверху чернеют, растрескиваются двумя скручивающимися створками и постепенно вытягиваются у некоторых видов до 10 см и более.
Все антоцеротовые отличаются дорсивентральным, лопастным вегетативным телом — слоевищем, или талломом. Слоевище состоит всего из нескольких слоев одинаковых, тонкостенных клеток. К краям оно обычно тоньше, и только у представителей рода дендроцерос (Dendroceros) посредине имеется многослойное утолщение, называемое жилкой. На нижней стороне таллома развиваются ризоиды с гладкими стенками, а в самом талломе, в его нижней части, у многих видов образуются межклеточные полости, открывающиеся на брюшной стороне щелеобразным отверстием (по-видимому, это редуцированное воздушное устьице). Полости заполнены слизью, которая окружает точку роста и предохраняет ее от высыхания. В полостях часто поселяются колонии сине-зеленой водоросли носток (Nostoc), выступающей как бы в роли симбионта с антоцеротовыми. Однако помещенные в культуру образцы антоцеротовых, у которых в слизистых полостях отсутствовали водоросли, продолжали нормально развиваться. Характерной особенностью антоцеротовых является наличие в клетках пластинчатого хлоропласта с пиреноидом (род антоцерос— Anthoceros) или нескольких более мелких хлоропластов с пиреноидами или без них (род мегацерос — Megaceros). У антоцеротовых органы полового размножения развиваются эндогенно в ткани слоевища. Гаметангии неравномерно разбросаны в средней части слоевища; часто женские и мужские оказываются на одном и том же растении. Антеридии располагаются по одному или группами в особых полостях — антеридиальных камерах, закрытых сверху одним или двумя слоями клеток. Из клеток ножки антеридия могут возникать дополнительные антеридии. Ко времени созревания антеридии окрашиваются в оранжевый цвет. Клетки, кроющие полость, вздуваются, затем кратерообразно разрываются и образуют низкую зубчатую обертку вокруг антеридиев, и сперматозоиды выходят наружу. Архегонии возникают также на спинной стороне, слоевища из поверхностных клеток, расположенных за верхушечной клеткой (слоевище при этом может продолжать свой рост). Архегонии у антоцеротовых не поднимаются над тканью слоевища и развиваются внутри его. Покровный слой архегония недостаточно отчетливо выделяется, поскольку его клетки срастаются с окружающими клетками гаметофита. Спорофит образуется из оплодотворенной яйцеклетки в ткани слоевища. По сравнению с низкоорганизованным, слабо расчлененным гаметофитом спорофит у антоцеротовых относительно высоко развит. У большинства представителей этого класса моховидных он состоит из длинной цилиндрической растрескивающейся коробочки, стопы и меристематического слоя между ними. У видов рода нототилас (Notothylas) имеется также рудиментарная ножка, а коробочка отличается ограниченным ростом.
Часть зародыша, из которого происходит спорогон, подразделяется на наружный слой — амфитеций (от греч. amphi — около и theke — вместилище) и внутренний — эндотеции (от греч. endon — внутри). Материнские клетки спор и элатер у антоцеротовых возникают из внутренней части амфитеция, так что споровый слой образует куполообразный свод над эндотецием, из которого развивается колонка, сходная с колонкой листостебельных мхов. Однако у видов рода нототилас колонка не образуется, и спорообразование у них происходит в эндотеции. Ткань слоевища над оплодотворенной яйцеклеткой разрастается в длинную и узкую цилиндрическую обертку вокруг молодого спорогона. У всех родов антоцеротовых, кроме нототиласа, спорогон, разрастаясь, прорывает ткань обертки. У подавляющего большинства антоцеротовых молодой спорогон зеленый: в клетках его стенок имеются хлоропласты с пиреноидами. Поэтому он даже способен к самостоятельному питанию, что доказано экспериментально. Те виды, которые в вегетативных клетках гаметофита имеют по одному хлоропласту, в клетках спорогона содержат по два хлоропласта. Виды с несколькими хлоропластами в вегетативных клетках имеют в клетках спорогона также большее их число (но только не удвоенное). И лишь у нототиласа в наружных клетках стенки спорогона хлоропласты отсутствуют. Развившийся спорогон (длиной от нескольких миллиметров до 13 см) у большинства родов обнаруживает сходство со «стручком». Ножка отсутствует, клубнеобразная стопа погружена в слоевище. Основание спорогона окружено оберткой из ткани гаметофита, так называемой вагинулой.
У многих родов стенка спорогона состоит из нескольких слоев клеток (например, у антоцероса). В ее наружном плотном слое расположены устьица, образованные двумя замыкающими клетками. Имеются и редуцированные формы спорогонов, у которых нет устьиц (нототилас). Центральная часть коробочки спорогона состоит из бесплодной колонки, но последняя обычно отсутствует у того же нототиласа. Между колонкой и стенкой коробочки развиваются тетрады спор и бесплодные нити — элатеры. Существует мнение, что колонка выполняет механическую функцию и служит также для проведения воды и питательных веществ. Образована она продольно вытянутыми клетками с узкими просветами. Элатеры у антоцеротовых могут быть одноклеточными или многоклеточными, иногда ветвистыми, часто коленчато согнутыми со спиральными утолщениями стенок или без них. Длительное время, примерно до образования тетрад спор, молодые и еще живые элатеры представляют собой нежные тонкостенные трубки с цитоплазмой, содержащей капельки масла и зерна крахмала, и выполняют роль питающих органов материнских клеток спор. Позднее на стенках элатер образуются спиральные утолщения. В зрелом спорогоне они становятся гигроскопичными и служат для разрыхления спор и разбрасывания их из коробочки. При основании спорогона, над его гаусториальной (всасывающей) частью, сохраняется меристематическая ткань. В результате деления ее клеток спорогон растет от основания вверх. Таким образом спорофит антоцеротовых, за исключением рода нототилас, обладает интеркалярным (вставочным) ростом, поэтому в верхней части спорогона споры созревают раньше, в то же время ближе к ее основанию споры еще лишь образуются. Таким I образом коробочка раскрывается постепенно сверху вниз двумя продольными щелями. Споры у антоцеротовых долго остаются соединенными в тетрады. Созревание спор антоцероса в условиях умеренной зоны происходит осенью. При прорастании споры образуется слабо развитая нитчатая протонема из одной или трех клеток; в роде дендродерос прорастание спор начинается еще в спорогоне.
Вегетативное размножение у антоцеротовых приурочено преимущественно к периодам с неблагоприятными для вегетации условиями (зимними, летними засушливыми). В такие периоды на краях или на нижней стороне слоевища формируются особые богатые запасными питательными веществами клубеньки, из которых могут вырасти новые растения. Некоторые многолетние антоцеротовые перезимовывают с помощью подземных частей слоевища, превращенных в клубеньки. К их числу относятся антоцеросы вильчатый и гималайский (Anthoceros dichotomus, A. himalayensis). Другие антоцеротовые принадлежат к однолетникам со слоевищем, разрушающимся после созревания и рассеивания спор. Относительно быстрое разрушение слоевищ и возобновление их преимущественно с помощью спор, по-видимому, обусловливают низкую конкурентную способность антоцеротовых. С этим связана и их экологическая приуроченность. Большинство видов является обитателями нарушенных и незадернованных местообитаний, поселяясь на залежах, по краю пашен, дорог и канав, вдоль берегов рек. Большей частью они выступают как пионеры при заселении обнаженной влажной почвы. В тропиках они часто переселяются с почвы на отмирающие дернины мхов, на полегшие травы, на кору гниющих стволов, на ветви и живые листья, где их слоевища могут успешно расти, избегая конкуренции с другими растениями.
В классе антоцеротовых выделяются либо одно семейство антоцеротовых (Anthocerotaсеае), либо два семейства: антоцеротовые и нототиласовые (Notothylaceae) с одним, и, как выше показано, своеобразным родом нототилас (Notothylas). Почти всеми признается самостоятельность рода антоцерос (Anthoceros), в широком его понимании, с включением в него аспиромитуса (Aspiromitus) и феоцероса (Phaeoceros), а также самостоятельность родов нототилас (Notothylas), дендроцерос (Dendroceros) и мегацерос (Megaceros). Основное число хромосом в классе 5—6. В классе антоцеротовых около 300 видов, широко распространенных главным образом в тропических и умеренно теплых областях земного шара. Около 200 видов относится к самому крупному роду антоцерос. Большинство видов антоцероса распространено в тропиках, но некоторые широко представлены в северных и южных умеренных широтах. Так, например, антоцерос гладкий (Anthoceros laevis) в Норвегии доходит до 60°30\’ с. ш. Самое же северное его местонахождение — в Исландии (карта 2). В России из всего класса антоцеротовых встречается только один род — антоцерос, представленный 3—4 видами.