Что такое сократимость мышцы
1. Роль мышечного сокращения в организации поведенческой деятельности человека. Классификация мышц и их функции. Виды и режимы мышечных сокращений.
2. Гладкие мышцы, их морфологические и физиологические особенности.
3. Одиночное мышечное сокращение, его фазы. Суммация сокращений.
4. Тетаническое сокращение, его виды. Оптимум и пессимум по Введенскому
5. Сократительный аппарат мышечного волокна. Механизм мышечного сокращения.
6. Синапс. Классификация. Особенности строения. Механизм передачи возбуждения в химическом синапсе. Свойства синапсов.
7. Медиатор. Виды медиаторов. Свойства медиаторов.
8. Электрические и тормозные синапсы. Особенности передачи сигнала.
9. Пути фармакологической регуляции синаптической передачи возбуждения.
1. Роль мышечного сокращения в организации поведенческой деятельности человека. Классификация мышц и их функции. Виды и режимы мышечных сокращений.
Общим свойством всего живого и основой активного поведения является движение.
Органом движения является мышечный аппарат, который включает 3 вида мышц: скелетные, гладкие и сердечную мышцы.
Они выполняют следующие функции:
1. Создание позы и удержание тела в пространстве, преодоление инерции.
1. Двигательная функция внутренних органов (моторная функция кишечника, сократительная функция сердца, обеспечение дыхания за счет сокращения дыхательных мышц).
2. Эффекторный механизм мыслительной (произносимая речь) и поведенческой деятельности.
3. Преобразование химической энергии макроэргических соединений в механическую, тепловую, электрическую энергию.
Составляют 35-40% массы тела, их количество достигает 600.
Состоят из пучков мышечных волокон, заключенных в общую соединительно-тканную оболочку.
Мышечное волокно – это гигантская, многоядерная мышечная клетка (диаметр от 1 до 100 мкм, длина от 5 до 400 мм), содержащая сотни миофибрилл, которые являются структурной единицей и представляют сократительный аппарат мышечного волокна. Миофибриллы включают актин и миозин.
Скелетные волокна подpазделяются на фазные волокна (они генерируют потенциал действия) и тонические (не способны генерировать распространяющееся возбуждение).
Фазные волокна делятся на быстрые волокна (белые, гликолитические) и медленные волокна (красные, окислительные).
Физические свойства скелетных мышц.
Растяжимость – это способность мышцы изменять свою длину под действием растягивающей силы.
Эластичность – способность мышцы принимать свою первоначальную длину после прекращения действия растягивающей или деформирующей силы.
Сила – определяется максимальным грузом, который мышца в состоянии поднять.
Способность совершать работу – определяется произведением массы поднятого груза на высоту подъема.
Физиологические свойства скелетных мышц: возбудимость, проводимость, сократимость, лабильность.
Для скелетной мышцы характерны три основных режима сокращения:
ИЗОТОНИЧЕСКИЙ – укорочение мышцы без изменения ее тонического напряжения (когда мышце не приходится перемещать груз, например, сокращение мышц языка).
ИЗОМЕТРИЧЕСКИЙ – длина мышечных волокон остается постоянной на фоне увеличения напряжения (попытка поднять непосильный груз)
АУКСОТОНИЧЕСКИЙ – изменение длины сопровождается изменением напряжения (работа мышцы при выполнении трудовых, спортивных и других двигательных актов).
Для скелетной мышцы характерны два вида сокращений:
ОДИНОЧНОЕ сокращение – возникает при действии одиночным стимулом (раздражителем) непосредственно на мышцу (прямое раздражение), или через иннервирующий ее двигательный нерв (непрямое).
ТЕТАНИЧЕСКОЕ (суммированное) сокращение – длительное сокращение мышцы в ответ на ритмическое раздражение.
(В естественных условиях к скелетной мышце из ЦНС поступают не одиночные импульсы, а серия импульсов, следующих друг за другом с определенными интервалами).
2. Гладкие мышцы, их морфологические и физиологические особенности
Гладкие мышцы находятся:
· во внутренних органах (пишеварительный тракт, мочевой пузырь);
· в сосудах, коже, глазе (мышцы радужной оболочки, цилиарная мышца).
Тонические – не способны развивать «быстрые» сокращения.
Фазно-тонические – способны быстро сокращаться и подразделяются на обладающие автоматией и не обладающие автоматией.
1. Образованы гладкомышечными клетками веретенообразной формы.
2. Хаотично расположены и окружены соединительной тканью (поэтому лишены поперечной исчерченности).
3. Контактируют друг с другом при помощи нексусов.
4. Сократительный аппарат представлен миофибриллами, состоящими в основном из актина. Миозин представлен только в дисперсной и агрегированной формах.
1. В основе сокращения – процесс превращения энергии АТФ в механическую энергию сокращения.
2. Сокращения медленные с использованием скользящего механизма.
3. Сокращение протекает с малыми энерготратами.
4. Обладают выраженной пластичностью (длительное сохранение измененной длины).
5. Обладают автоматией.
1. Быстрое и сильное растяжение гладких мышц.
2. Химические вещества (особенно гормоны и медиаторы, к которым гладкие мышцы обладают высокой чувствительностью).
Особенности электрических процессов.
1. Потенциал покоя в гладких мышцах меньше, чем в скелетных.
В клетках не обладающих автоматией он стабилен и =– 60–70 мВ.
В клетках, обладающих автоматией, он неустойчивый с колебаниями от –30 до –70 мВ.
Потенциал действия имеет длительный латентный период.
Ниже, чем в скелетных мышцах.
Бывает двух типов: пикоподобная форма и форма «плато».
Связан с повышением проницаемости для ионов Са.
Несколько опережает сокращение.
2. Проведение возбуждения возникает, если приложенный стимул одновременно возбуждает некоторое минимальное количество мышечных клеток.
Может распространяться на соседние мышечные волокна (из-за малого сопротивления в области контактов) распространяется лишь на определенное расстояние, которое зависит от силы раздражителя скорость значительно меньше, чем в скелетной мышце и составляет от 2 до 15 см/с.
3. Одиночное мышечное сокращение, его фазы. Суммация сокращений
Одиночное мышечное сокращение продолжается около 100 мс и развивается по фазам:
Латентный (скрытый) период продолжается до 3 мс и представляет время от начала действия раздражителя до начала видимого ответа (сокращения) мышцы.
Фаза сокращения продолжается 40-50 мс характеризуется укорочением длины мышечного волокна, что связано с увеличением концентрации Сa 2+ в протофибриллярных пространствах и образованием актин-миозиновых связей.
Фаза расслабления продолжается 50-60 мс характеризуется увеличением (восстановлением) длины волокна. Возникает при снижении концентрации Ca 2+ в протофибриллярных пространствах и ослаблением актин-миозиновых связей.
Если на мышцу наносятся два и более раздражений с интервалом менее продолжительности одиночного сокращения, но более продолжительности рефрактрного периода ПД, то происходит суммация сокращений, в результате которой сократительный эффект усиливается.
Существует два типа суммации: частичная и полная
Частичная (или неполная) суммация возникает, если
· интервал между раздражениями меньше продолжительности одиночного мышечного сокращения;
· больше продолжительности фазы сокращения, т.е. если второе раздражение попадает в фазу расслабления.
В результате амплитуда мышечного сокращения возрастает с образованием двух вершин.
Полная суммация возникает, если:
· интервал между раздражениями меньше продолжительности фазы сокращения, но больше продолжительности рефрактерного периода;
· второе раздражение попадает в фазу сокращения.
В результате амплитуда мышечного сокращения изменяется (увеличивается или уменьшается относительно одиночного сокращения) с образованием одной вершины
Увеличение или уменьшение амплитуды связано с изменением возбудимости в процессе возбуждения и зависит от того в какую фазу измененной возбудимости наносится следующее раздражение.
Известно, что в период формирования пикового потенциала возбудимость ткани снижена (фаза абсолютной и фаза относительной рефрактерности). Поэтому, если следующее раздражение будет наноситься в этот период, то амплитуда мышечного сокращения будет снижена.
Период возбуждения в скелетной мышце завершается следовой деполяризацией, продолжающейся от 20 до 40 мс.
В этот период возбудимость, а, следовательно, и сократимость повышена. Поэтому, если следующее раздражение будет приходиться на этот период, то амплитуда мышечного сокращения будет возрастать (тем больше, чем больше повышена возбудимость).
3. Тетаническое сокращение, его виды. Оптимум и пессимум по Введенскому
Различают два вида тетануса: зубчатый и гладкий.
В их основе лежат механизмы частичной или полной суммации.
Вид тетанического сокращения определяется Механическим состоянием мышцы в момент повторного возбуждения. Состоянием возбудимости мышцы в момент повторного возбуждения.
Зубчатый тетанус развивается на ряд последовательных раздражений, интервал между которыми больше продолжительности фазы сокращения, но меньше продолжительности одиночного мышечного сокращения (интервал от 100 до 50 мс при частоте раздражений от 10 до 20 Гц).
При этом каждое новое сокращение формируется на фоне не завершившегося расслабления мышцы, образуя новые вершины последующих сокращений («зубцы»). Высота суммарного сокращения зависит от ритма и силы раздражений и определяется исходным уровнем формирования каждого следующего сокращения (чем выше уровень, тем больше амплитуда).
В начале фазы расслабления этот уровень выше, чем в конце.
Гладкий тетанус развивается на ряд последовательных раздражений, интервал между которыми меньше длительности фазы сокращения, но больше продолжительности потенциала действия (интервал от 50 до 5 мс при частоте 20 до 200 Гц).
Каждое новое сокращение формируется на фоне не завершившегося сокращения мышцы, образуя единую, гладкую вершину. Ее высота определяется уровнем измененной возбудимости в процессе возбуждения.
Если каждый следующий раздражитель попадает в фазу экзальтации (повышенной возбудимости), то амплитуда сокращения будет большой.
Если импульсы попадают в период сниженной возбудимости (относительная рефрактерность), то амплитуда будет снижена.
Явление изменения амплитуды в зависимости от возбудимости мышцы объяснил H.Е.Введенский, введя понятие оптимума и пессимума.
Оптимальная частота – максимальная частота раздражений, при которой возникает максимальная амплитуда тетанического ответа.
Пессимум – снижение амплитуды тетанического сокращения при увеличении частоты раздражений (выше оптимальной величины).
Пессимальная частота – максимальная частота (сверх оптимальной), при которой возникает минимальная амплитуда тетанического ответа.
4. Сократительный аппарат мышечного волокна. Механизм мышечного сокращения
Структурной единицей мышечного волокна являются миофибриллы.
Они разделены на чередующиеся участки (диски), которые обладают различными оптическими свойствами.
Диски, обладающие двойным лучепреломлением, получили название анизотропные (А) диски.
Диски, которые не обладают двойным лучепреломлением, названы изотропные (I) диски.
Анизотропные диски в обыкновенном свете выглядят темными и состоят из двух темных полосок, разделенных светлой «H» полоской.
Изотропные диски в обыкновенном свете выглядят светлыми и в середине имеют темную «Z» полоску.
Z полоска – эта тонкая мембрана, которая является продолжением поверхностной мембраны вглубь мышечного волокна.
Она выполняет опорную функцию, поскольку через ее поры проходят протофибриллы.
В зоне Z мембраны также находятся триады или Т-системы триады представляют выпячивания плазматической мембраны с образованием поперечных трубочек в виде ярусов и цисцерн.
Они предсталяют саркоплазматический ретикулум, который содержит высокую концентрацию ионов Ca.
При возбуждении Z мембраны кальций по концентрационному градиенту выходит из саркоплазматического ретикулума в протофибриллярное пространство, вызывая процесс сокращения Активная реабсорбция ионов Са в саркоплазматический ретикулум за счет работы Са-насоса, приводит к расслаблению мышечного волокна.
Структурной единицей миофибриллы являются протофибриллы
Протофибриллы включают белковые нити актина и миозина, а также белки тропонин и тропомиозин.
Нити миозина – это толстые и короткие нити, которые входят только в состав анизотропного диска.
Нити актина – это тонкие и длинные нити, входящие в состав как изотропного, так и анизотропного дисков. Они вставлены между нитями миозина. От них свободна только H-полоска анизотропного диска.
Процесс сокращения происходит в результате скольжения нитей актина относительно нитей миозина, который запускается накоплением Са, при этом образуются актино-миозиновые комплексы (мостики) и нити актина вдвигаются в промежутки между нитями миозина.
Нити актина сближаются друг с другом.
Ширина H-полоски и анизотропных дисков уменьшается, изотропный диск своего не изменяет своего размера.
Механизм мышечного сокращения и расслабления.
2. Возникновение потенциала действие.
3. Проведение возбуждения вдоль клеточной мембраны до Z мембраны, а далее вглубь волокна по трубочкам саркоплазматического ретикулума.
4. Освобождение Са из триад.
5. Диффузия Са к протофибриллам.
6. Взаимодействие Са с тропонином.
7. Конформационное изменение комплекса тропомиозин-тропонин.
8. Освобождение активных центров актина.
9. Присоединение актина к миозину.
10. В присутствии белка актомиозина распад АТФ с освобождением энергии.
11. Скольжение нитей актина относительно миозина.
12. Укорочение миофибриллы.
13. Активация кальциевого насоса.
15. Понижение концентрации свободных ионов Са в саркоплазме.
16. Разрушение актин-миозиновых комплексов.
17. Обратное скольжение нитей актина относительно миозина.
18. Увеличение (восстановление) миофибриллы.
5. Синапс. Классификация. Особенности строения. Механизм передачи возбуждения в химическом синапсе. Свойства синапсов
Синапс (соединять, смыкать, связывать) – это структурное образование, которое обеспечивает переход возбуждения с нервного волокна на инервируемую клетку.
Классификация и особенности строения по учебнику.
Механизм синоптической передачи возбуждения.
1. Деполяризация (возбуждение) пресинаптической мембраны.
2. Изменение проницаемости для ионов кальция.
3. Ионы кальция или его ионизированные комплексы по концентрационному градиенту поступают в нервное окончание (антагонистами кальция являются ионы магния и токсины ботулинуса).
4. Уменьшение электростатических влияний (одноименных зарядов) между пресинаптической мембраной и везикулами.
5. Приближение и слияние везикул с пресинаптической мембраной.
6. Изменение поверхностного натяжения везикул.
8. Выход медиатора в синоптическую щель.
9. Медиатор (возбуждающий в нервно-мышечном синапсе: ацетилхолин) диффундирует через синоптическую щель к рецепторам постсинаптической мембраны.
10. Ацетилхолин вступает во взаимодействие с холинорецепторами (обладают избирательной чувствительностью к ацетилхолину).
11. При одновременном участии ионов кальция и макроэргического фосфата происходят конформационные изменения белковых молекул рецептора.
12. Открываются каналы постсинаптической мембраны для Na или Са.
13. Ионы Na по концентрационному градиенту поступают внутрь воспринимающей возбуждение клетки.
14. Развивается деполяризация – возбуждающий постсинаптический потенциал, который носит местный характер, по форме и свойствам напоминает локальный ответ (не подчиняется закону «все или ничего» и способен суммироваться).
15. Суммация возбуждающих постсинаптических потенциалов
16. Потенциал концевой пластинки.
17. Когда он достигает определенной (критической величины) возникают местные токи между возбужденными участками постсинаптической мембраны и невозбужденными участками прилегающей к ней обычной (электровозбудимой) мембраной.
18. На прилегающем участке электровозбудимой мембраны возникает потенциал действия.
Свойства химических синапсов.
1. Нервно-химический механизм передачи возбуждения (передача возбуждения осуществляется с помощью специфического химического вещества – медиатора, который выделяется нервным окончанием и количество которого пропорционально частоте приходящей нервной импульсации).
2. Принцип Дейла (во всех синапсах, образованных нервными окончаниями одного нейрона, выделяется только один вид медиатора – либо возбуждающий, либо тормозный).
3. Одностороннее проведение возбуждения (возбуждение передается только в одном направлении – от пресинаптической мембраны к постсинаптической мембране).
4. Синаптическая задержка (скорость проведения возбуждения в синапсе значительно медленнее, чем в нервном и мышечном волокне).
5. Низкая функциональная лабильность синапса.
6. Трансформация ритма возбуждения (при большой частоте нервных импульсов в пресинаптичоском окончании происходит уменьшение частоты возбуждений в постсинаптических образованиях).
7. Высокая утомляемость (временная потеря работоспособности в результате несоответствия синтеза и расхода медиатора).
Вопросы для самостоятельной подготовки (по учебнику).
1. Медиатор. Виды медиаторов. Свойства медиаторов.
2. Электрические и тормозные синапсы. Особенности передачи сигнала.
3. Пути фармакологической регуляции синаптической передачи возбуждения.
Биомеханические свойства скелетных мышц человека
В третьей лекции по дисциплине «Биомеханика мышц» для студентов НГУ им. П.Ф.Лесгафта рассматриваются биомеханические свойства скелетных мышц человека: сократимость, жесткость, вязкость, прочность, релаксация. Рассмотрена трехкомпонентная модель мышцы.
Лекция 3
Биомеханические свойства скелетных мышц человека
Анализируя предмет биомеханики, А.А. Ухтомский (1927) указывал: «Биомеханика изучает ту же систему нервно-мышечных приборов как рабочую машину, то есть задается вопросом, каким образом полученная механическая энергия движения и напряжения может приобрести определенное рабочее применение» (С. 141). Начиная с этой лекции, мы будем рассматривать именно этот аспект деятельности мышц.
3.1. Биомеханические свойства мышц
Биомеханические свойства скелетных мышц – это характеристики, которые регистрируют при механическом воздействии на мышцу.
Следует отметить, что в условиях живого организма изучение биомеханических свойств мышц крайне затруднено. В этой лекции, кроме биомеханических свойств мышц, приводятся данные о свойствах сухожилий и связок.
К биомеханическим свойствам мышц относятся:
Сократимость
Сократимость – способность мышцы укорачиваться при возбуждении, в результате чего возникает сила тяги.
В первой лекции было подробно рассмотрено строение первичного сократительного элемента мышцы – саркомера. В 1966 году А. Гордон, А. Хаксли и Ф. Джулиан провели специальные исследования, позволившие установить зависимость силы, развиваемой саркомером, от его длины. Одно из предположений, касающихся механизма скольжения филаментов, заключалось в том, что каждый поперечный мостик (миозиновая головка) действует подобно независимому генератору силы. Поэтому уровень силы, развиваемой во время сокращения, должен зависеть от количества одновременных взаимодействий между толстыми и тонкими филаментами. Это предположение подтвердилось. Действительно, существуют критические значения длины саркомера, при которых развиваемая им сила падает до нуля (рис.3.1).
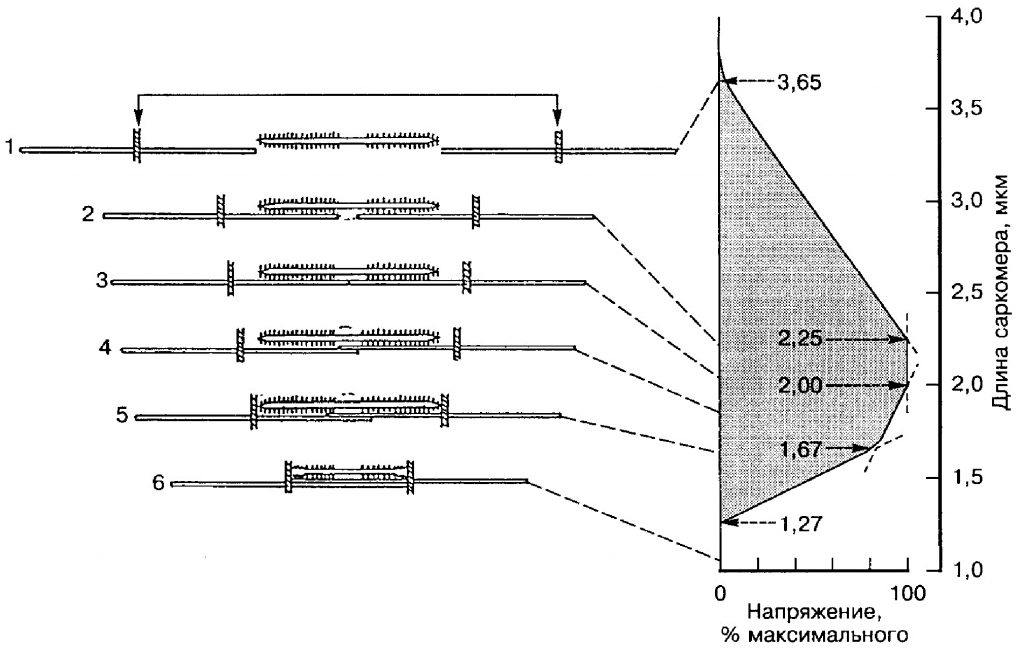
Рис. 3.1. Схема, иллюстрирующая зависимость между степенью перекрытия толстых и тонких филаментов и силой, развиваемой саркомером (по: A.M. Gordon, A.F. Huxley. F.J. Julian, 1966)
Первое критическое значение длины саркомера равно 1,27 мкм. Оно соответствует максимальному укорочению мышцы. В этом состоянии мышцы регулярность расположения толстого и тонкого филаментов нарушается, они искривляются. Поэтому количество одновременных взаимодействий между филаментами резко уменьшается. Сила падает до нуля. Второе критическое значение длины саркомера равно 3,65 мкм. Оно соответствует максимальному удлинению мышцы. При максимальном растяжении саркомера перекрытия толстых и тонких филаментов нет, поэтому сила уменьшается до нуля. Если длина саркомера находится в интервале от 1,27 мкм до 3,65 мкм, значение силы отличается от нуля. Максимальная сила, которую способен развить саркомер, соответствует значениям его длины – от 1,67 до 2,25 мкм.
Жесткость
Жесткость материала – характеристика тела, отражающая его сопротивление изменению формы при деформирующих воздействиях (В.Б. Коренберг, 2004). Чем больше жесткость тела, тем меньше оно деформируется под воздействием силы. Закон Гука гласит, что сила упругости, возникающая при растяжении или сжатии тела, пропорциональна его удлинению.
Жесткость материала характеризуется коэффициентом жесткости (k). Единица измерения жесткости тела – Н/м. Жесткость линейной упругой системы, например, пружины, есть величина постоянная на всем участке деформации.
В отличие от пружины, мышца представляет собой систему с нелинейными свойствами. Это связано с тем, что структура мышцы очень сложна. Поэтому для мышцы зависимость силы от удлинения будет отлична от закона Гука. Возникающая в мышце сила упругости не пропорциональна удлинению. Вначале мышца растягивается легко, а затем даже для небольшого ее растяжения необходимо прикладывать все большую силу. Поэтому часто мышцу сравнивают с трикотажным шарфом, который вначале легко растягивается, а затем становится практически нерастяжимым. Иными словами, жесткость мышцы с ее удлинением возрастает. Из этого следует, что мышца представляет собой систему, обладающую переменной жесткостью. В этом случае коэффициент жесткости k равен первой производной силы по деформации материала. Установлено, что жесткость активной мышцы в 4-5 раз больше жесткости пассивной мышцы. В табл. 3.1. представлены значения коэффициентов жесткости мышц-сгибателей стопы у представителей разных видов спорта.
Таблица 3.1 Значения коэффициента жесткости мышц-сгибателей стопы у представителей различных видов спорта
(по: А.С. Аруину, В.М. Зациорскому, Л.М. Райцину, 1977)
Вязкость
Вязкость – свойство жидкостей, газов и «пластических» тел оказывать неинерционное сопротивление перемещению одной их части относительно другой (смещение смежных слоев). При этом часть механической энергии переходит в другие виды, главным образом в тепло (В.Б. Коренберг, 1999).
Это свойство сократительного аппарата мышцы вызывает потери энергии при мышечном сокращении, идущие на преодоление вязкого трения. Предполагается, что трение возникает между толстыми и тонкими филаментами при сокращении мышцы. Кроме того, трение возникает между возбужденными и невозбужденными мышечными волокнами. Это связано с тем, что соседние мышечные волокна «связаны» посредством эндомизия. Поэтому, если возбуждены все мышечные волокна, трение должно быть меньше. Показано, что при сильном возбуждении мышцы, ее вязкость резко уменьшается (Г.В. Васюков,1967).
Если абсолютно упругое тело (например, пружину) вначале растянуть, а затем – снять деформирующую нагрузку, то кривая «удлинение – сила» будет идентичной во время обеих фаз. Если же мы имеем дело с упруговязким материалом (мышцей), кривые окажутся неидентичными. При нагрузке (растягивании мышцы) зависимость «удлинение – сила» соответствует кривой 1. Рис.3.2.
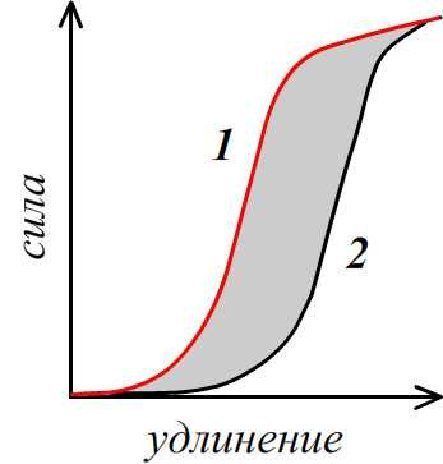
Рис. 3.2. Зависимость «удлинение – сила» при растягивании (кривая 1) и укорочении мышцы (кривая 2)
При укорочении мышцы зависимость «удлинение – сила» соответствует кривой 2. Кривые 1 и 2 образуют «петлю гистерезиса». Площадь фигуры, заключенной между кривыми 1 и 2, отражает потери энергии на трение. Мышца, обладающая большей вязкостью, будет характеризоваться большей площадью «петли гистерезиса». Вы знаете, что при выполнении физических упражнений температура мышц повышается. Повышение температуры мышц связано с наличием у мышц вязкости. Результатом наличия вязкости происходят потери энергии мышечного сокращения на трение. Разогрев мышц (разминка) приводит к тому, что вязкость мышц уменьшается.
Прочность
Прочностью материала называют его способность сопротивляться разрушению под действием внешних сил (И.Ф. Образцов с соавт., 1988).
Значительно снижает прочность связок и сухожилий иммобилизация. И, наоборот, при исследовании животных была найдена связь между уровнем физической активности и прочностью сухожилий и связок. Показано, что в подавляющем большинстве случаев прочность сухожилий более высока, чем прочность их прикрепления к костям. Поэтому при травмах сухожилий они не разрываются, а отрываются от места прикрепления. Следует учитывать также, что в процессе тренировок прочность сухожилий и связок увеличивается сравнительно медленно. При форсированном развитии скоростно-силовых качеств мышц может возникнуть несоответствие между возросшими скоростно-силовыми возможностями мышечного аппарата и недостаточной прочностью сухожилий и связок. Это грозит потенциальными травмами (А.С. Аруин, В.М. Зациорский, В.Н. Селуянов, 1981).
Релаксация
Релаксация мышц – свойство, проявляющееся в уменьшении с течением времени силы тяги при постоянной длине.
Для оценки релаксации используют показатель – время релаксации, то есть отрезок времени, в течение которого натяжение мышцы уменьшается в е раз от первоначального значения. Многочисленными исследованиями установлено, что высота выпрыгивания вверх с места зависит от длительности паузы между приседанием и отталкиванием. Чем больше эта пауза (изометрический режим работы мышц), тем меньше сила их тяги и, как следствие, высота выпрыгивания, табл. 3.2. Таким образом, релаксация мышц приводит к уменьшению высоты выпрыгивания.
Таблица 3.2 Влияние паузы на высоту прыжка с места (n = 31) (по: А.С. Аруин, В.М. Зациорский, Л.М. Райцин, 1977)